

Las semillas de malezas en la cosecha de soja: ¿Amenaza para los sistemas agrícolas?
Weed seeds in soybean crop: threat to farming systems?
 Luz Fabiola García fabiolagarcia@hotmail.com
Carolina Waigand carolina.waigand@uner.edu.ar
Fabián Abel Ayala fabian.4y4l4@gmail.com
María del Rosario Baigorria rosariobai93@gmail.com
Luz Fabiola García fabiolagarcia@hotmail.com
Carolina Waigand carolina.waigand@uner.edu.ar
Fabián Abel Ayala fabian.4y4l4@gmail.com
María del Rosario Baigorria rosariobai93@gmail.com
Revista FAVE Sección Ciencias Agrarias
Universidad Nacional del Litoral, Argentina
ISSN: 2346-9129
ISSN-e: 2346-9129
Periodicidad: Semestral
núm. 23, e0022, 2024
Recepción: 10 Abril 2023
Aprobación: 07 Marzo 2024
Autor de correspondencia: silvia.ledesma@uner.edu.ar

Resumen: Las malezas constituyen adversidades importantes para el desarrollo, cosecha y rendimiento de los cultivos. Las prácticas agronómicas modifican las comunidades de malezas, según las características de las especies: producción de semillas, fenología, morfología, resistencia a herbicidas. El objetivo de este estudio fue caracterizar la comunidad de especies cuyas semillas están presentes en granos de soja cosechados en el centro-oeste de la provincia de Entre Ríos. Se tomaron muestras de soja de las campañas 2020 y 2021 en cuatro Departamentos de la provincia de Entre Ríos (Argentina). Se describió la fracción de Otras Semillas de cada muestra por su tamaño y riqueza específica; se calculó la abundancia relativa y frecuencia de especies por muestra; y la diversidad específica de la comunidad. Se estableció la similitud florística entre campañas y entre Departamentos y se sometieron las muestras a un análisis multivariado para detectar agrupamientos. No hubo diferencias significativas en tamaño y riqueza de la fracción entre campañas ni entre Departamentos. Se determinó una única comunidad representativa de ambas campañas con marcada dominancia de Amaranthus hybridus L. y Echinochloa crus-galli (L.) de reconocida habilidad competitiva y resistencia múltiple a herbicidas. La comunidad de Otras Semillas en soja es una potencial amenaza para los sistemas agrícolas, por la dispersión de semillas de especies resistentes hacia nuevas áreas; y por su perpetuación en el banco de semillas del suelo.
Palabras clave: semillas, malezas, soja, amenaza.
Abstract: Weeds constitute important adversities for crop development, harvest and yield. Agronomic practices modify weed communities according to species characteristics: seed production, phenology, morphology, and herbicide resistance. The objective of this study was to characterize the community of species whose seeds are present in soybeans harvested in the center-west of Entre Ríos province. Soybean samples were taken from the 2020 and 2021 seasons in four departments of Entre Ríos (Argentina). The Other Seeds fraction of each sample was described by size and specific richness; the relative abundance and frequency of species per sample was calculated; and the specific diversity of the community. The floristic similarity between campaigns and between Departments was established and the samples were subjected to multivariate analysis to detect clusters. There were no significant differences in size and richness of the fraction between campaigns or between Departments. A single representative community was determined for both campaigns with a marked dominance of Amaranthus hybridus L. and Echinochloa crus-galli (L.), of recognized competitive ability and multiple resistance to herbicides. The community of other seeds in soybean is a potential threat to agricultural systems, by the dispersal of seeds of resistant species to new areas, and by their perpetuation in the soil seed bank.
Keywords: seeds, weeds, soybean, threat.
INTRODUCCIÓN
Las malezas constituyen adversidades importantes de los sistemas agroproductivos, dificultando el desarrollo de los cultivos y eventualmente la cosecha y disminuyendo el rendimiento y la calidad de los granos (Schwartz-Lázaro et al., 2022). Las prácticas agronómicas (sistema de labranza, rotaciones de cultivos, aplicaciones de herbicidas) y los efectos ambientales (variaciones estacionales e interanuales del clima, la erosión del suelo, cambio climático) modifican las poblaciones de malezas perjudicando a algunas y favoreciendo a otras (Ghersa y León 1999; Martínez-Ghersa et al., 2000; Tuesca et al., 2001; Poggio, 2012; Scursoni et al., 2014; Kruk, 2015). En Argentina, el cultivo de soja basado en siembra directa produjo cambios en la flora de malezas incluyendo desaparición de algunas especies, aparición de otras, disminución de la riqueza, incremento en la abundancia relativa de gramíneas y especies con semillas transportadas por el viento; y disminución de latifoliadas anuales (Puricelli y Tuesca, 2005; Papa y Tuesca, 2008).
La propagación por semillas constituye una vía exitosa para muchas especies (Nisensohn et al., 2011; Uztarroz et al., 2016; Alshallash, 2018; Chauhan et al., 2019; Niño-Hernández et al., 2020, Alonso y Bornard, 2021) que en ocasiones se potencia con el desarrollo de nuevos biotipos adaptados a variables ecológicas y de manejo (Romagnoli et al., 2013; Yanniccari, 2014).
A la vez, la dispersión a través de las maquinarias involucradas en la cosecha ha sido señalada por su importancia en distintos lugares del mundo (Barroso et al., 2006; 2012); verificándose lo mismo también en la región agrícola núcleo de Argentina (Tourn et al., 2018). Y a esto puede sumarse que la dispersión a través de la contaminación de las semillas puede incrementarse cuando los agricultores guardan sus propias semillas en lugar de utilizar semillas certificadas (Rao et al., 2017).
El objetivo de este estudio fue caracterizar la comunidad de especies cuyas semillas están presentes en granos de soja cosechados en el centro-oeste de la provincia de Entre Ríos.
MATERIALES Y MÉTODOS
Área de Estudio
El área de estudio comprendió cuatro Departamentos del centro oeste de la provincia de Entre Ríos: Paraná, Diamante, Victoria, Nogoyá. Los tres primeros conforman la Zona Agroeconómica (ZA) Paraná, que Engler et al. (2008) describen como una zona dominada por un paisaje suavemente ondulado (“casi peniplanicie”); pero donde también se registran áreas de pendientes pronunciadas. Se trata de una región afectada principalmente por procesos de degradación de suelos por erosión hídrica; perteneciendo en su mayoría a Argiudoles y Hapludoles; con algunas zonas arenosas (Udisamentes). Por otro lado, la ZA Nogoyá, presenta un ambiente donde se acentúan las características pampeanas, con predominio de suelos vérticos; y una configuración de peniplanicie con valles aluviales en torno a los arroyos. Se trata de suelos con aptitud agrícola con distintos niveles de limitaciones. Ambas zonas presentan aptitud agrícola, con soja y trigo como cultivos principales. El clima es templado húmedo, con una media anual de 1100 – 1200 mm de precipitaciones.
Condición hídrica
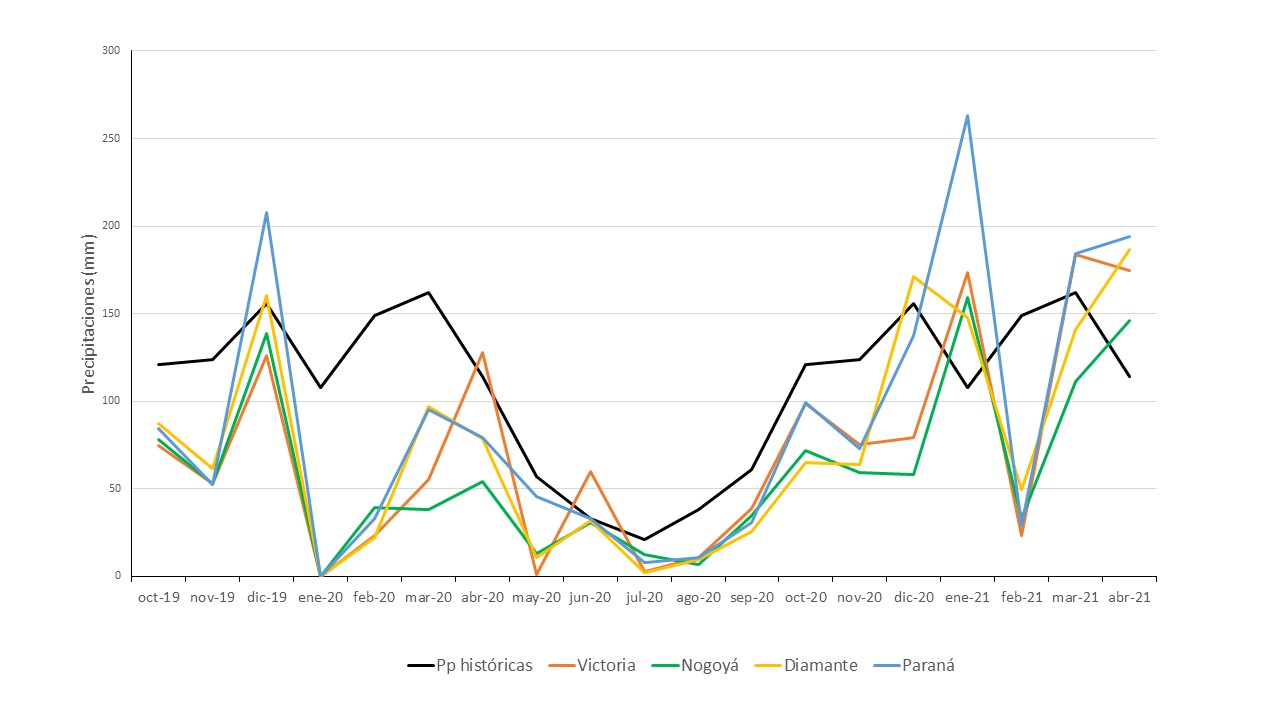
Se realizó la caracterización de las precipitaciones ocurridas durante el período octubre 2019 a abril 2021 para contextualizar el desarrollo del cultivo y la comunidad de malezas. Los datos de precipitaciones se consultaron en los registros de la Bolsa de Cereales de Entre Ríos.
Obtención de las muestras
En dos campañas de producción sucesivas (2019-2020 y 2020-2021) se tomaron muestras de granos de soja en cuatro centros de acopio de los Departamentos Nogoyá, Victoria, Diamante y Paraná (Entre Ríos, Argentina); completando 17 muestras de la campaña 2019-2020 (S2020) y 15 de la campaña 2020-2021 (S2021) de un kg cada una. Se extrajeron inmediatamente después la descarga desde los acoplados de transporte y previo a cualquier tipo de manipulación o limpieza, a los efectos de garantizar que estas muestras representen las condiciones de infestación de malezas resultantes del ciclo a campo.
Caracterización de la comunidad de malezas en muestras de soja
En el Laboratorio de Análisis de Semillas FCA UNER (RNCyFS 1281/I), se procedió al análisis de las muestras siguiendo los conceptos y lineamientos generales de los Análisis de Pureza Físico-Botánica y de Análisis de Semillas en Número (ISTA, 2023). A partir de cada muestra se separó la fracción de Otras Semillas (OS) en forma manual, por observación directa y/o con lupa binocular. ISTA (2023) define a la fracción de OS como aquella que “incluye unidades de semillas de cualquier especie de planta que no sean de la semilla pura del cultivo” presentes en la muestra de trabajo. Posteriormente se procedió a la identificación de las especies de la fracción de OS en base a bibliografía disponible, bases de datos online y archivo de semillas del Laboratorio; y se contabilizó el número total de unidades de semilla cada una de las especies de OS identificadas. En la fracción de Otras Semillas se determinó: Tamaño de la fracción (TOS), dado por el número total de unidades de Otras Semillas presentes por muestra; Número de unidades de semilla pura por especie; y Riqueza Específica (RE): número total de especies de OS identificadas por muestra. Además, se caracterizó fitosociológicamente a la comunidad de especies mediante la determinación de Abundancia relativa por especie: relación entre el número de unidades de semilla de cada especie y el tamaño de la fracción de OS de la muestra, expresado en porcentaje; Frecuencia: porcentaje de aparición de cada especie en relación al total de muestras por campaña y Diversidad específica (H) medida por el Índice de Shannon-Weaver (1949):
Donde: S: riqueza especifica; π.: promedio de abundancia relativa por muestra de cada especie
Y se complementó con el cálculo de diversidad máxima para esa comunidad: Hmax = lnS; indicando la condición en la cual existe un aporte homogéneo por parte de cada especie (Pla, 2006).
Las comunidades de la fracción de OS de cada campaña se compararon mediante el Índice de Similitud florística de Sorensen (S):
Donde a: suma de los valores de abundancia relativa de las especies de la comunidad 1; b: suma de los valores de abundancia relativa de las especies de la comunidad 2; y c: suma de los menores valores de abundancia relativa de las especies en común entre las dos comunidades.
Análisis estadístico
Se utilizó el ANAVA Simple (α=0,05), tomando como variables de respuesta TOS y RE por muestra; y Departamento de origen y año de producción como variables explicativas. Con el test de Tuckey (p.<0,05) se determinó la existencia de diferencias significativas. Se realizaron transformaciones matemáticas de variables a los efectos de cumplir con los supuestos de normalidad exigidos por los métodos estadísticos, a partir de los valores de curtosis exhibidos por las variables en estudio.
Con la finalidad de detectar agrupamientos de las muestras caracterizadas por el número de unidades de semilla pura por especie de OS, fueron sometidas al análisis multivariado utilizando un método de conglomeración jerárquico aglomerativo. Este método inicia poniendo cada muestra en un conglomerado separado. Luego, los conglomerados son unidos, dos cada vez, hasta que el número de conglomerados es reducido a un objetivo deseado. En cada etapa, los conglomerados son unidos en pares por cercanía, utilizando en este estudio el método de encadenamiento completo (vecino más lejano), tomando la distancia euclidiana como medida de similitud.
El software utilizado fue Infostat Versión 2013 (Di Rienzo et al., 2013).
RESULTADOS
Condición hídrica
Las dos campañas presentaron períodos deficitarios de precipitaciones en comparación con la media histórica para la zona (Figura 1), en los cuatro Departamentos. En S2020 el registro promedio de los cuatro Departamentos estuvo 55,4% por debajo de la media en la segunda mitad del ciclo del cultivo enero2020 – abril2021, mientras que la campaña 2020-2021 se inició con déficit de humedad y luego se desarrolló con recuperación de valores de humedad para completarse con la lluvia caída entre noviembre 2020 y abril 2021 alcanzando un total apenas 11% menor al valor histórico (Figura 1).
Caracterización de la fracción de Otras Semillas
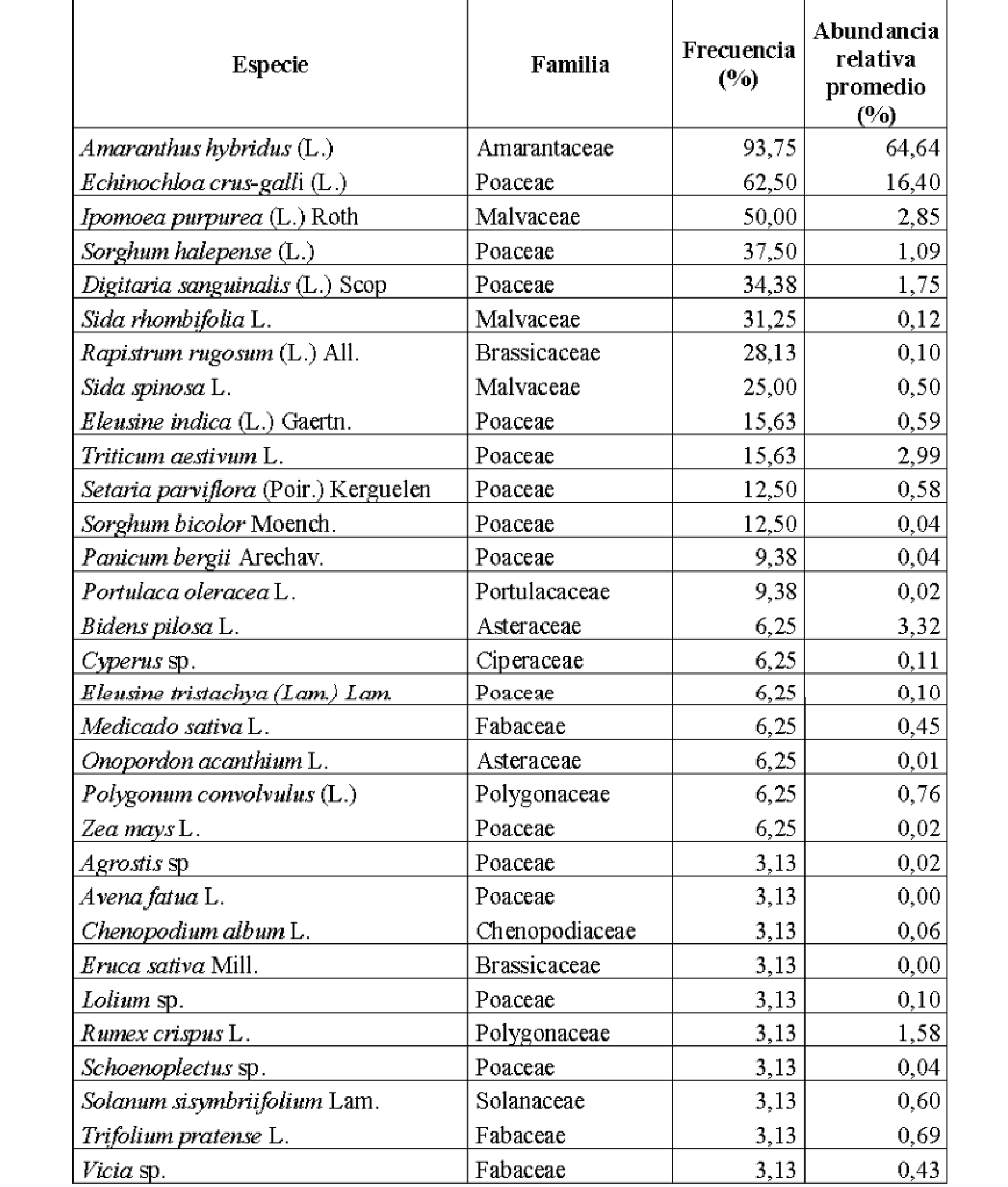
El TOS en S2020 varió entre 63 y 102795 unidades por muestra (Tabla 1) con un promedio de 7117 unidades de semilla por muestra (Tabla 2). Por su parte, la RE de OS por muestra en S2020 varió entre 1 y 9 especies (Tabla 1), registrándose 21 especies en el total de muestras de ese ciclo y alcanzándose una RE promedio de seis especies de OS por muestra (Tabla 2).
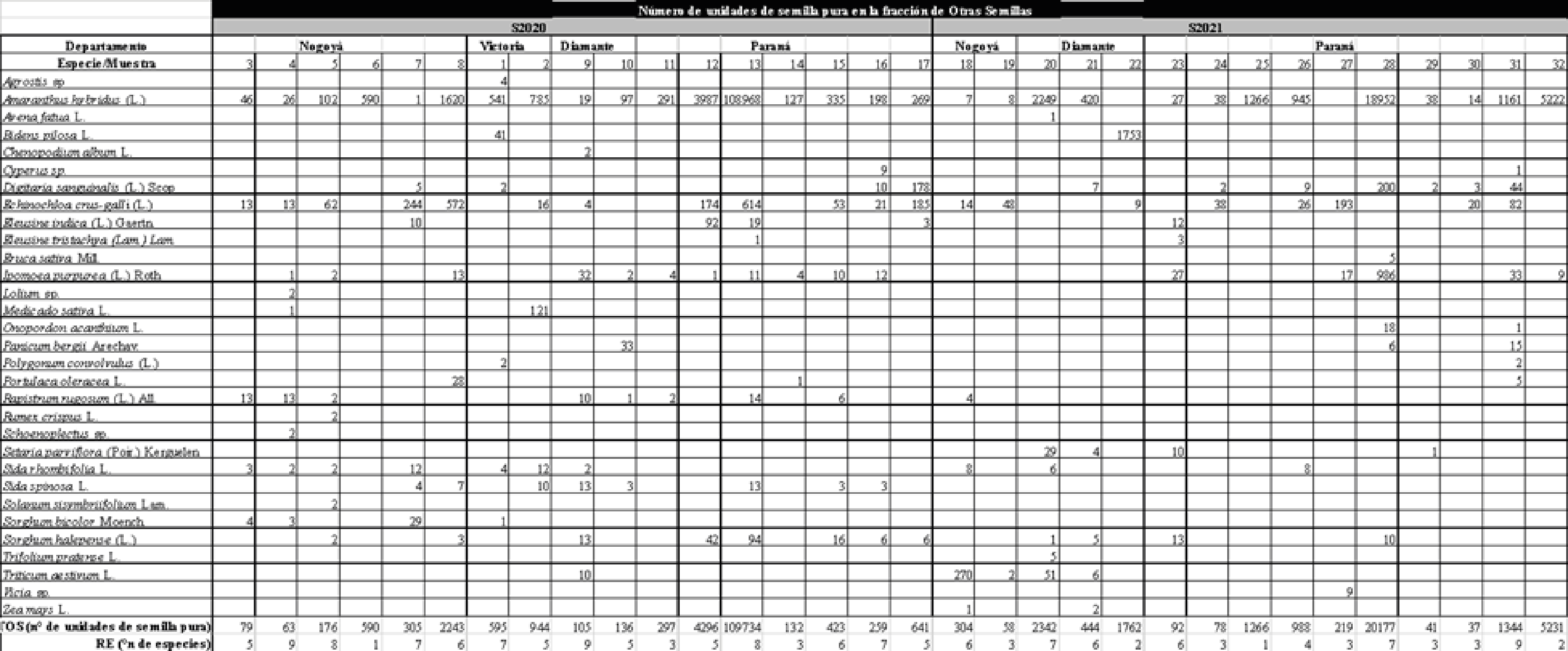
En S2021 el TOS varió entre un mínimo de uno y un máximo de 20177 unidades (Tabla 1), obteniéndose un promedio de 2311 unidades de OS por muestra (Tabla 2). En total se identificaron 24 especies de OS en S2021 (Tabla 1), mientras que la RE por muestra alcanzó un mínimo de una especie y un máximo de nueve (Tabla 1) con un promedio de 4,6 especies por muestra (Tabla 2).
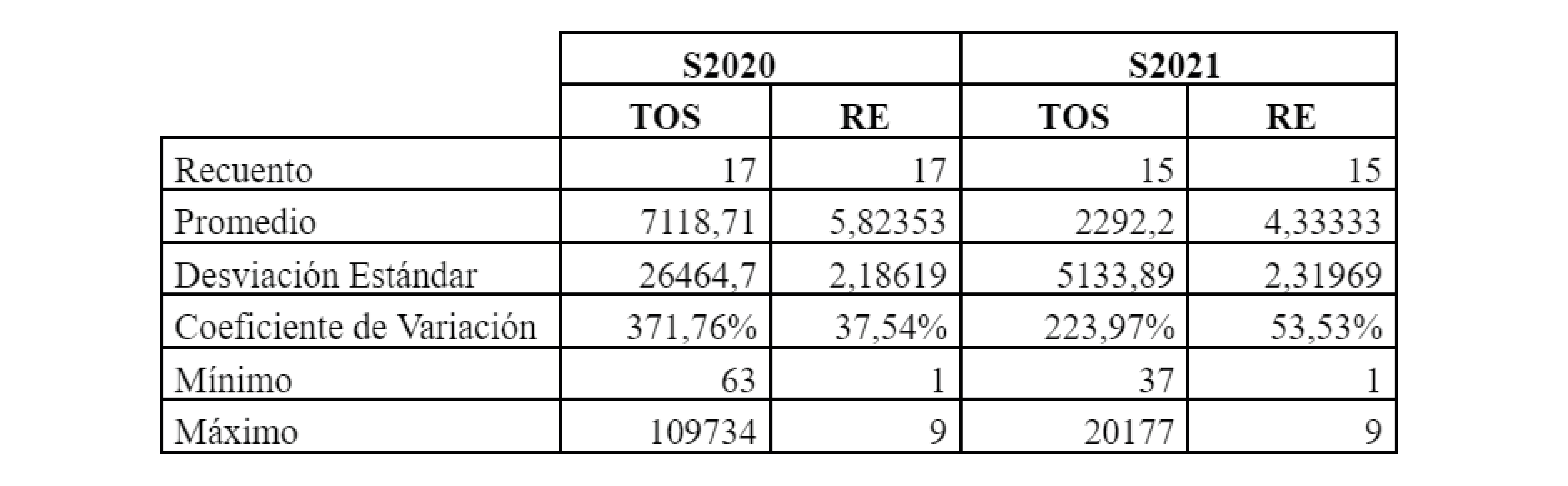
Los valores de TOS y RE no resultaron significativamente distintos (p<0,05) entre años de producción ni tampoco se observaron diferencias significativas de estas variables entre muestras tomadas en distintos departamentos en cada uno de los ciclos productivos.
Características fitosociológicas por año de cosecha
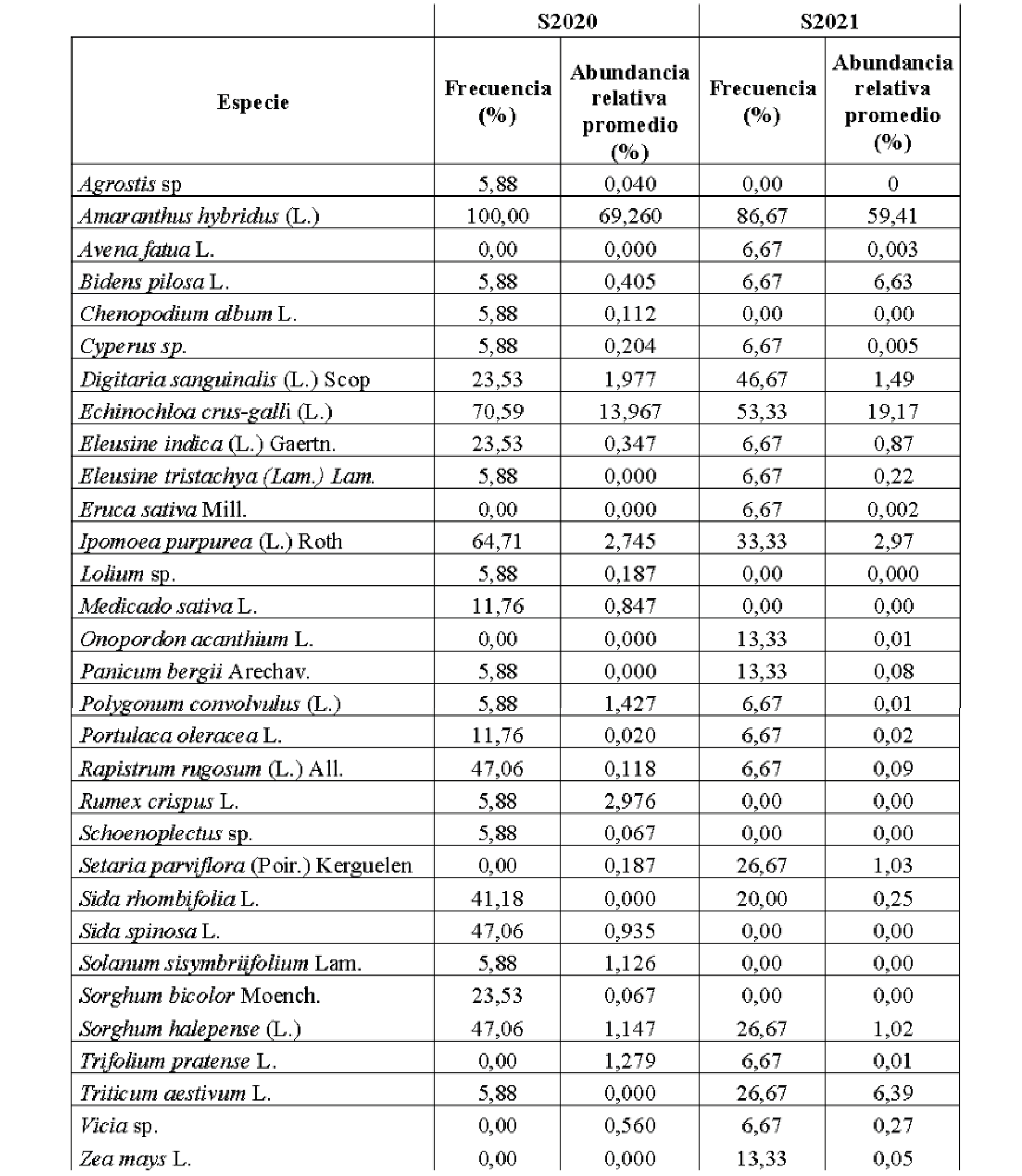
En S2020, A. hybridus fue la especie de mayor abundancia relativa promedio (69,27%) presentándose en el 100% de las muestras y le siguió E. crus-galli (capín), cuya abundancia relativa por muestra fue del 14,21% en promedio (Tabla 3); registrándose en el 70,6% de las muestras. D. sanguinalis, R. rugosum y S. halepense se presentaron en más del 40% de las muestras, al igual que dos cultivos voluntarios (M. sativa y T. aestivum). El resto de las especies no superaron el 2,96% de abundancia relativa en promedio por muestra (Tabla 3).
En S2021 también A. hybridus fue la más frecuente (75%) y de mayor abundancia relativa promedio (47,9%); y E. crus-galli se encontró en el 40% de las muestras, con 16,07% de abundancia relativa promedio. I. purpurea apareció en el 45% de las muestras con una abundancia relativa promedio de 11,85% (Tabla 3). Todas las demás especies de OS de la S2021 registraron abundancia relativa menor al 5%, y se registraron seis especies cultivadas: T. aestivum, O. sativa, S. bicolor, Vicia sp., Z. mays y T. pratense.
Comparación entre Departamentos de origen y ciclos de producción
El índice de similitud de Sorensen se calculó en 70,47% indicando que la fracción de OS de las campañas de soja 2020 y 2021 constituyen una misma comunidad representativa de ambas ZA.
Por su parte, los dendrogramas basados en TOS por muestra y abundancia relativa por especie, demostraron que no hubo agrupamientos de las muestras en cada año por Departamento de origen ni por campaña de producción (Figura 2 a y b), corroborando la determinación de una sola comunidad de OS.
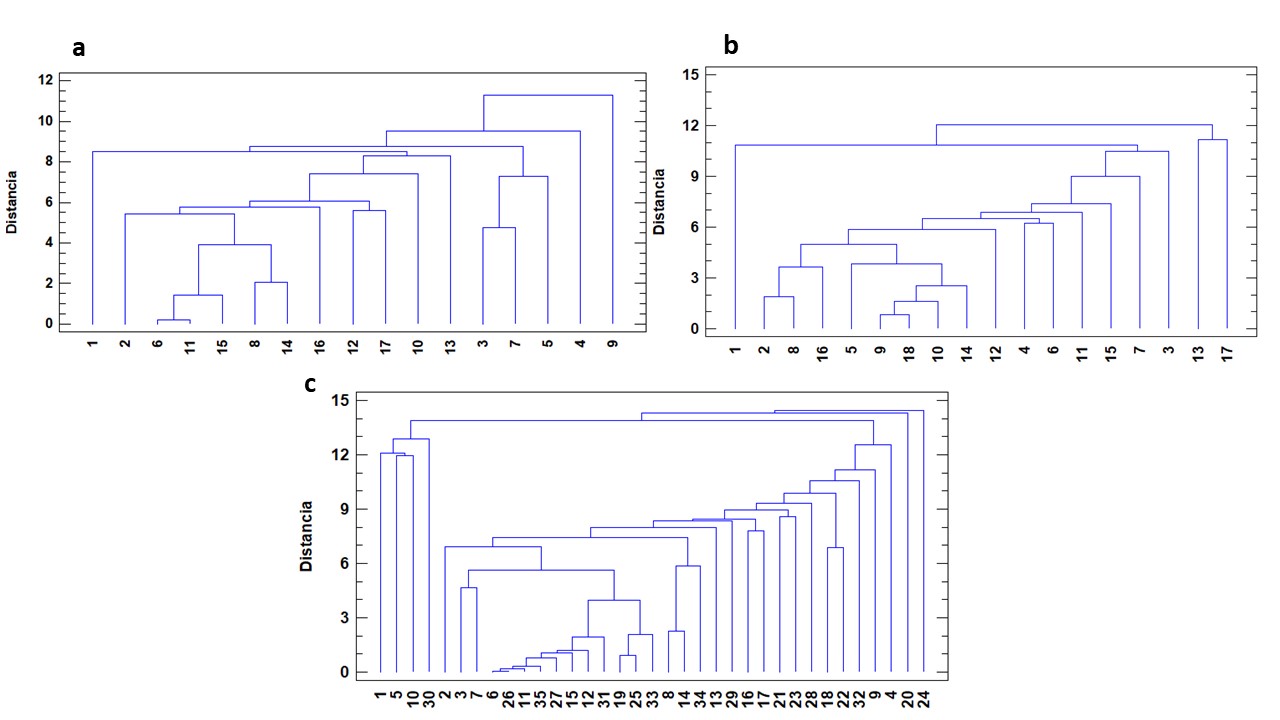
El dendrograma que involucra la totalidad de las muestras (Figura 2c) presenta un conglomerado constituido por el 31,4% de las muestras; las que provienen en proporciones similares de ambas campañas y pertenecen a las dos ZA. Se trata de muestras que presentaron una elevada proporción de A. hybridus en la fracción de OS (mayor a 80%). Por otro lado, se observaron otros dos pequeños grupos que incluyen tan solo 2 y 3 muestras. En esta disposición queda reflejada la gran heterogeneidad observada tanto en el TOS como en la composición florística y la abundancia relativa de las especies en la fracción de OS en ambas campañas y zonas de producción.
En consecuencia, la fracción de OS de cosecha de soja de las ZA Paraná y Nogoyá (Entre Ríos); sin discriminar año de cosecha puede describirse con un TOS promedio de 4835,66 + 19417,7 unidades de semilla; y una RE que promedió 5,31+2,44 especies por muestra.
Fitosociología de la fracción de Otras Semillas en cosecha de soja de las ZA Paraná y Nogoyá
En la fracción de OS de las ZA Paraná y Nogoyá (E. Ríos) se determinó la presencia de una comunidad constituida por 31 especies, y dominada ampliamente por A. hybridus, cuya abundancia relativa promedio fue de 64,64%; con una frecuencia de 93,75% (Tabla 4). La segunda especie en abundancia relativa promedio fue E. crus-galli; con 16,40%; y una frecuencia de 62,5%. Otras seis especies se presentaron en más del 25% de las muestras, y 13 alcanzaron frecuencia entre 5 y 15%. El resto solamente se presentó en una muestra (Tabla 4). El 18,75% de las especies alcanzaron valores de abundancia relativa promedio entre 1 y 10%, mientras que la mayoría presentó valores de abundancia relativa por debajo de 1% (Tabla 4).
La comunidad incluyó 11 familias botánicas (Figura 3a) donde Poaceae fue la más representada (47% del total de especies). Tanto Malvaceae como Fabaceae aportaron 3 especies cada una (Figura 3a), aunque las Fabaceae encontradas no se consideran malezas, sino que resultan de la resiembra voluntaria de cultivos comerciales (M. sativa, T. pratense y Vicia sp.).
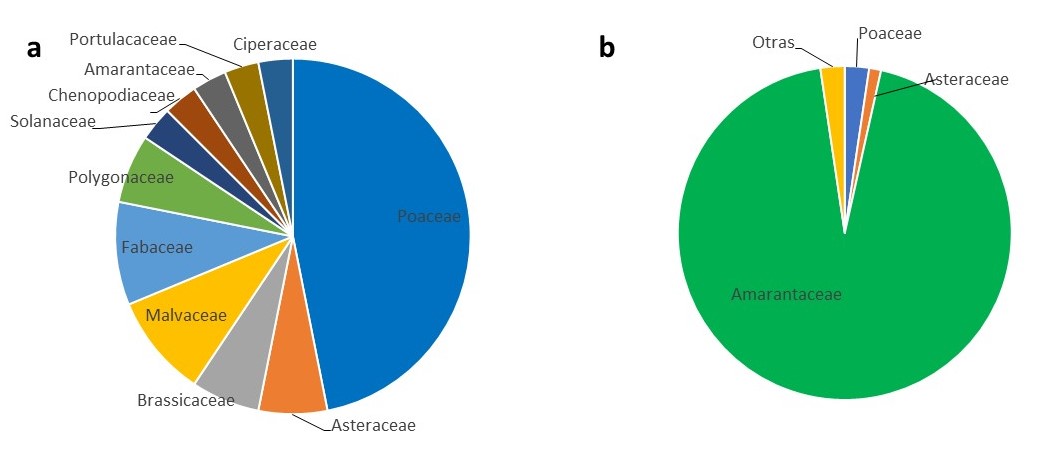
Amarantaceae aportó la mayor cantidad de unidades de semilla (95% del total de unidades de semilla de OS), todas de A. hybridus. Le siguieron Poaceae; Convolvulaceae y Asteraceae (Figura 3b); aunque entre las tres sólo concentraron el 4,5% del total de unidades colectadas. El resto de las familias aportaron en conjunto un 1% del total de unidades de OS (Figura 3b).
Finalmente, se determinó una diversidad calculada en H´= 1,547; habiéndose estimado un máximo de Hmax = 3,367 en caso de que existiera una equitativad óptima en abundancia relativa por especie.
DISCUSIÓN
La comunidad de OS en soja de las campañas 2020 y 2021 del centro-oeste de Entre Ríos tuvo una RE promedio de 5,3 especies, un valor bajo y coincidente con lo descripto por Poggio (2012) quien atribuye la baja riqueza de especies a la aplicación de siembra directa y el uso de cultivos transgénicos y glifosato en la Región Pampeana. En este sentido, de la Fuente et al. (2006) advierten sobre la pérdida de recursos genéticos y su impacto sobre los servicios ecosistémicos en el mundo, como consecuencia de las prácticas agrícolas.
El tamaño de la fracción de OS por muestra promedió las 4836 unidades. Forcella et al. (1996) reportan que en los granos de mayor tamaño como soja o maíz suelen presentarse menor cantidad de unidades de OS, en comparación con lo que ocurre con trigo o canola, por ejemplo; atribuyéndolo a la eliminación de semillas extrañas durante la trilla en la máquina cosechadora en acuerdo a lo explicado por Schwartz-Lazaro et al. (2022). En este aspecto, Tourn et al. (2018) al registrar la presencia de malezas en el 100% de las cosechadoras que evaluaron en la Región Pampeana (Argentina) demostraron que esta maquinaria representa una de las vías de mayor importancia para la diseminación de estas plagas.
Por otro lado, el TOS depende también de la densidad de individuos de malezas que alcancen la etapa de semillazón en coincidencia con el cultivo, lo que les permite también alimentar el banco de semillas del suelo (Foresto et al., 2021).
El presente estudio detectó una única comunidad de malezas en la fracción de OS de la soja cosechada en cuatro departamentos de Entre Ríos en dos campañas sucesivas. Dado que las precipitaciones fueron similares en toda el área, se infiere una uniformidad del manejo aplicado al cultivo de soja, especialmente en lo relacionado con el control de malezas.
Por otro lado, la composición florística de la comunidad de OS puede estar influenciada por condiciones propias de la cosecha. Wilson et al. (2016) menciona que las malezas de porte más alto y con semillas retenidas en la parte superior, son las que luego se detectan en el análisis de semillas. Además, es probable encontrar también semillas de cultivos “voluntarios”, tal como se dio en este estudio; donde se registró la presencia de semillas de trigo, raigrás, maíz y alfalfa; con abundancia relativa promedio menor al 5%.
Tourn et al. (2018) registraron variabilidad de densidades y flora de semillas en cosechadoras, destacándose la abundancia de Lolium sp.; Avena fatua y A. palmeri., malezas de porte alto y con resistencia a herbicidas.
Sangoy Puntín et al. (2021) registraron una Riqueza Específica de 35 especies en cultivo en pie de soja, con lo cual se podría suponer que la mayoría de las malezas no llegan a contaminar con sus semillas el grano que se cosecha, aunque es probable que sí generen una retroalimentación del banco de semillas del suelo.
La comunidad de malezas determinada en este trabajo tuvo a A. hybridus como especie dominante. Las especies del género Amaranthus poseen diversas habilidades competitivas como: adaptación a stress térmico e hídrico, alta producción de semillas (entre 37.000 y 600.000 semillas por planta), alto vigor de sus semillas; viabilidad en el suelo por largos períodos; fotoblastismo negativo, longevidad, germinación en profundidad, entre otras (De Marco et al., 2014; Niño-Hernandez et al., 2020); sumadas al desarrollo de resistencia a herbicidas (Schwartz-Lázaro et al., 2017; Gaweda et al., 2020; Aulakh et al., 2021). E. crusgalli, I. purpurea, S. halepense podrían identificarse como especies acompañantes en la comunidad, por sus valores intermedios de abundancia y frecuencia. Schwartz-Lazaro et al. (2022) señalan la presencia de A. hybridus, Ipomoea sp. y Ambrosia sp., en soja, y destacan su capacidad de retener semillas en planta en la madurez del cultivo propiciando su presencia en la cosecha. Asimismo, Sangoy Puntín et al. (2021), indican que A. hybridus, se presenta como una de las especies más abundantes en lotes de soja y en la biomasa cosechada, aún con utilización de herbicidas. Por otro lado, en sistemas que no usaron control químico, esto autores citan a E. crusgalli, S. halepense, D. sanguinalis como malezas presentes en mayor proporción.
En este trabajo, la comunidad de OS estuvo dominada por A. hybridus y E. crus-galli, dos especies con resistencia múltiple a herbicidas en Argentina y el mundo (Chahal et al., 2015; Scursoni et al., 2022); y del total de especies de OS registradas, el 70% ha sido reportada en la lista de la la International Herbicide Resistant Weed Database (Heap, s.f.) por presentar uno o más tipos de resistencia a herbicidas en el mundo En particular, la Red de Manejo de Plagas de la Asociación Argentina de Productores de Siembra Directa (AAPRESID, s.f.) incluye en su listado de especies con alertas de resistencia a herbicidas en Argentina a ocho de las que formaron parte de la comunidad de OS de soja determinada en este estudio: A. hybridus, A. fatua, D. sanguinalis, E. indica, Lolium sp., R. sativus, R. rugosum, S. halepense. Esto constituye una alerta para las investigaciones en la región, dado que la detección temprana es fundamental en la selección y adopción de medidas de manejo de los biotipos resistentes. En este sentido, Díaz et al. (2009) sugieren aplicar técnicas de detección de resistencia a nivel de semillas de malezas, por lo cual resultaría de gran utilidad el análisis de pureza físico-botánica y de semillas en número de los lotes de soja cosechados.
Las características de la comunidad de OS advierten sobre la probabilidad de difusión de resistencia a herbicidas a nuevas áreas geográficas mediante la maquinaria de cosecha (Tourn et al., 2018); y sobre el depósito de las mismas en el banco de semillas del suelo (Brainard et al., 2008). En la región del Espinal de Entre Ríos, se verificó presencia de semillas de malezas con resistencia a herbicidas en suelos de bosques nativos aledaños a campos agrícolas (Sione et al., 2015), indicando el potencial de infestación sobre ecosistemas naturales
CONCLUSIONES
La comunidad de Otras Semillas de la cosecha de soja en las Zonas Agroeconómicas Paraná y Nogoyá (Entre Ríos) advierte sobre la presencia y potencial difusión de especies de alta importancia como malezas de la agricultura. Si bien es una comunidad de baja riqueza específica, está fuertemente dominada por Amaranthus hybridus y Echinochloa colonum, especies con alta capacidad de producción de semillas en una fenología coincidente con el cultivo de soja; morfología que facilita la dispersión a través de los equipos de cosecha y trilla; y resistencia múltiple a herbicidas.
En general, la comunidad de OS se compone en un 78% de especies con algún tipo de resistencia a herbicidas, incluyendo un 30% que ya ha sido reportadas en Argentina.
En consecuencia, la comunidad de OS identificada constituye una potencial amenaza para los agroecosistemas tanto por la dispersión de semillas de especies resistentes hacia nuevas áreas; como por su perpetuación en el suelo. Resultan prioritarios los estudios para la detección temprana de biotipos resistentes en la región; en pos de la selección y adopción de prácticas sustentables de manejo y control de malezas en agroecosistemas de la región.
Agradecimientos
Este trabajo fue realizado en el marco del PID UNER 2211, financiado por la Universidad Nacional de Entre Ríos. Los autores agradecen la colaboración de COOPAR; LAR, Dellizzotti Hnos. SRL y de los Ings. Agrs. Carlos E. Toledo† y Hernán Pintos.
REFERENCIAS
Alonso, C. S. y Bornand, C.L. (2021). Dinámica de emergencia de malezas primavera-estivales en Villa Mercedes, San Luis, Argentina. Revista Científica y Técnica Agropecuaria, Agroindustrial y Ambiental, 8, 2. http://servicios.ingenieria.unlz.edu.ar:8080/ojs/index.php/agrarias/article/view/87
Alshallash. K. (2018). Germination of weed species (Avena fatua, Bromus catharticus, Chenopodium album and Phalaris minor) with implications for their dispersal and control. Annals of Agricultural Sciences, 63(1), 91-97.
Asociación Argentina de Productores de Siembra Directa (AAPRESID). (s.f.). Mapa de malezas resistentes. Recuperado de: https://www.aapresid.org.ar/rem/malezas#monitoreo
Aulakh, J., Chahal, P., Kumar, V., Price, A., y Guillard, K. (2021). Multiple herbicide-resistant Palmer amaranth (Amaranthus palmeri) in Connecticut: Confirmation and response to POST herbicides. Weed Technology, 35(3), 457-463, https://doi.org/10.1017/wet.2021.6
Barroso, J.; Navarrete, L.; Sánchez del Arco M.J.; Fernandez-Quintanilla, C.; Lutman, P.J.; Perry, N.H. y Hull, R.I. (2006). Dispersal of Avena fatua and Avena sterilis patches by natural dissemination, soil tillage and combine harvesters. Weed Research, 46(2), 118-128, https://doi.org/10.1111/j.1365-3180.2006.00500.x
Barroso, J., Andújar D., San Martín, C., Fernandez-Quintanilla C., y Dorado J. (2012). Johnson - grass (Sorghum halepense) Seed Dispersal in Corn Crops under Mediterranean Conditions. Weed Science 60, 34-41.
Brainard, D. C., Bellinder, R. R., Hahn, R. R., y Shah, D. A. (2008) Crop rotation, cover crop and weed management effects on weed seed banks and yields in snap bean, sweet corn and cabbage. Weed Science, 56, 434-441.
Chauhan, B.S., Ali, H.H. y Florentine, S. (2019). Seed germination ecology of Bidens pilosa and its implications for weed management. Science Reports 9, 16004. https://doi.org/10.1038/s41598-019-52620-9
Chahal, P.S., Aulakh, J.S., Jugulam, M., y Jhala, A. (2015). Herbicide-Resistant Palmer amaranth (Amaranthus palmeri S. Wats.) in the United States. En Price, A., Kelton, J., y Sarunaite, L. (Eds.) Mechanisms of Resistance, Impact, and m Management. IntechOpen. http://dx.doi.org/10.5772/61512
de la Fuente, E.B., Suarez, S.A., y Ghersa, C. M. (2006). Soybean weed community composition and richness between 1995 and 2003 in the Rolling Pampas (Argentina). Agriculture, Ecosystems and Environment, 115, 229–236, doi:10.1016/j.agee.2006.01.009
De Marco, N.; Sabattini, R.; Sione, S.; Ledesma, S.; Anglada, M.; Olea, I.; Sabaté, S.; Vincinguerra, H. y Lovato Echeverría, R. (2014). Echinochola colona (L.) Link; Echinochloa crusgalli (L.) P. Beav. Echinochloacruspavonis (Kunth) Schult. En: Fernandez, O; Leguizamón, E. & Acciaresi, H. (Eds.) Malezas e invasoras III. (1ª. ed.; pp. 329-347). Ediuns.
Di Rienzo J.A., Casanoves F., Balzarini M.G., González L., Tablada M., y Robledo C.W. (2013). InfoStat (Internet). Versión 2013. Córdoba: Grupo InfoStat; 2013. http://www.infostat.com.ar
Díaz, J., Espinoza, N., & Galdames, R. (2009). Test de germinación in vitro para detectar malezas resistentes a herbicidas. Serie Actas - Instituto de Investigaciones Agropecuarias N° 44. https://hdl.handle.net/20.500.14001/8578
Engler, P., Rodríguez, M., Cancio, R., Handloser, M., y Vera, L.M. (2008). Zonas AgroEconómicas Homogéneas de Entre Ríos. Descripción Ambienta, socioeconómica y productiva. Recuperado de: https://inta.gob.ar/sites/default/files/script-tmp-inta_zonas_agroeconmicas_homogneas_entre_ros.pdf
Forcella, F., Peterson, D. H., y Barbour, J. C. (1996). Timing and measurement of weed seed shed in corn (Zea mays). Weed Technology, 10, 535–543, doi: 10.1017/S0890037X00040409
Foresto, E., Amuchástegui, M.A., Nuñez, O, y Zorza, E. (2021). Comportamiento del banco de semilla de malezas en un sistema agrícola-ganaderointervenido por diferentes tipos de labranzas. Ciencia e Interculturalidad, 28(01), 151-165. https://doi.org/10.5377/10.5377/rci.v28i01.11466
Gaweda, D., Haliniarz, M., Bronowicka-Mielniczuk, U., y Łukasz, J. (2020). Weed Infestation and Health of the Soybean Crop Depending on Cropping System and Tillage System. Agriculture, 10, 208, doi:10.3390/agriculture10060208.
Ghersa, C. M., y León, R.J.C. (1999). Successional changes in agroecosystems of the rolling pampas. En L.R. Walker (ed.). Ecosystems of the world. Ecosystems of disturbed ground (pp. 487-502). Elsevier.
Heap, I. (s.f.). The International Herbicide-Resistant Weed Database. Online. Recuperado de: https://www.weedscience.org/Pages/Species.aspx
International Rules for Seed Testing Association. (2023). Full Issue i–19-10 (308)
Kruk B.C. (2015). Disminución de la emergencia de malezas en diferentes escenarios agrícolas bajo siembra directa. Rev. Agronomía & Ambiente 35(2), 179-190.
Martínez-Ghersa, M.A., Ghersa C.M., y Satorre, E.H. (2000). Coevolution of agricultural systems and their weed companions: implications for research. Field Crops Research 67, 181-190.
Niño-Hernandez, J.C., Felipe Moreno, D., Ruiz-Berrío, H.D., Balaguera-López, H.E., y Magnitskiy, S. (2020). Luz, giberelinas y profundidad de siembra inciden sobre la germinación de semillas de Amaranthushybridus L. Revista U.D.C.A Actualidad & Divulgación Científica, 23, 2, http://doi.org/10.31910/rudca.v23.n2.2020.1545.
Nisensohn, L. A., Tuesca, D., y Papa, J. C. (2011). Diferencias en la susceptibilidad al glifosato en plantas de Conyzabonariensis (L.) Cronquist y Conyza sumatrensis (Retz) con distinto grado de desarrollo. Revista Para Mejorar la Producción, 46, 105-108.
Papa, J.C., y Tuesca, D.H. (2008). Malezas del Cultivo de Soja. CREA Producción de Soja. pp 71-78
Pla, L. (2006). Biodiversidad: inferencia basada en el Ííndice de Shannon y la riqueza. INCI 3, 8
Poggio, S. (2012). Cambios florísticos en comunidades de malezas: un marco conceptual basado en reglas de ensamblaje. Ecología Austral, 22, 150-158.
Puricelli, E., y Tuesca, D. (2005). Efecto del sistema de labranza sobre la dinámica de la comunidad de malezas en trigo y en barbechos de secuencias de cultivos resistentes a glifosato. AgriScientia, 22(2). https://doi.org/10.31047/1668.298x.v22.n2.2682
Rao, A.N., Brainard, D.C., Kumar, V., Ladha, J.K., y Johnson, D.E. (2017). Chapter Two - Preventive Weed Management in Direct-Seeded Rice: Targeting the Weed Seedbank. Advances in Agronomy 144: 45-142. https://doi.org/10.1016/bs.agron.2017.02.002
Romagnoli, M. V., Tuesca, D., y Permingeat, H. R. (2013). Caracterización de la resistencia de Amaranthus quitensis a tres familias de herbicidas. Ecología Austral, 23(2), 119–125.
Sangoy Puntin, N., Poggio, S.L., y Coll, L. (2021). Productividad de soja de segunda y comunidades de malezas en sistemas de intensificación sustentable y de transición ecológica. En: ¿Cómo integramos la soja en los sistemas productivos? Avances y perspectivas en Entre Ríos (Ed. Santos, D.) Serie de Extensión INTA Paraná - Nº88.
Schwartz-Lazaro, L. M., Green, J. K., y Norsworthy, J. K. (2017). Seed retention of Palmer amaranth (Amaranthus palmeri) and barnyardgrass (Echinochloa crus-galli) in soybean. Weed Technology, 31, 617–622. https://doi.org/10.1017/wet.2017.25
Schwartz-Lazaro, L., LaBiche, G. y Copes, J.T. (2022). Detection of Foreign Material in Soybean (Glycine max) Grain. Frontiers in Agronomy, 4, 868573. doi: 10.3389/fagro.2022.868573.
Scursoni, J.A., Gigòn, R., Martìn, A. N., Vigna, M., Leguizamòn, C., y López Source, R. (2014). Changes in Weed Communities of Spring Wheat Crops of Buenos Aires Province. Weed Science, 62(1), 51-62. http://dx.doi.org/10.1614/WS-D-12-00141.1
Scursoni, J. A., Tuesca, D., Balassone, F., Morello, J. P., Herrera, D. M., Lescano, M. C., ... y Vila-Aiub, M. M. (2022). Response of smooth pigweed (Amaranthus hybridus) accessions from Argentina to herbicides from multiple sites of action. Weed Technology, 36(3), 384-389. https://doi.org/10.1017/wet.2022.9
Shannon, C.E., y Weaver, W. (1949) The mathematical theory of communication. University of Illinois Press. Urbana, IL, EEUU. 144 pp.
Sione, S. M. J., Ledesma, S.G., Rosenberger, L. G., Galliussi, R., y Sabattini, R. A. (2015). Banco de semillas del suelo, en relación a dos estados sucesionales del bosque nativo en Entre Ríos. Quebracho-Revista de Ciencias Forestales, 23(2), 62-76.
Tourn, S.N., Diez de Ulzurrum, P., Exilart, A., Lasaga, R., y Platz, J.P. (2018). Presencia y abundancia de semillas de malezas en máquinas cosechadoras. Malezas, 1, 6-11. https://doi.org/10.15258/istarules.2023.F
Tuesca, D., Puricelli, E. y Papa, J.C. (2001). A long-term study of weed flora shifts in different tillage systems. Weed Research, 41, 369-382.
Uztarroz, D., Kruk, B.C., Satorre, E. H., y Ghersa, C. M. (2016). Dormancy, germination and emergence of Urochloa panicoides regulated by temperature. Weed Research, 56(1), 59-68. https://doi.org/10.25260/EA.13.23.2.0.1176
Wilson, C. E., Castro, K. L., Thurston, G. B., y Sissons, A. (2016). Pathway risk analysis of weed seeds in imported grain: A Canadian perspective. NeoBiota, 30, 49–74. doi: 10.3897/neobiota.30.7502
Yannicari, M. (2014). Estudio fisiológico y genético de biotipos de Loliumperenne L. resistentes a glifosato. [Tesis Doctoral, Universidad Nacional de La Plata]. https://doi.org/10.35537/10915/37620
Notas de autor
silvia.ledesma@uner.edu.ar

