

Artículos
Efecto de la salinidad sobre el crecimiento de plantines de Cucurbita maxima Duch. x Cucurbita moschata Duch
Effect of salinity on the growth of seedlings of Cucurbita maxima Duch. x Cucurbita moschata Duch
 Franco Ángel Rossi
Gastón Devecchi
Alvaro Codaro
Gabriel Céccoli
Carlos Alberto Bouzo
Franco Ángel Rossi
Gastón Devecchi
Alvaro Codaro
Gabriel Céccoli
Carlos Alberto Bouzo
Revista FAVE Sección Ciencias Agrarias
Universidad Nacional del Litoral, Argentina
ISSN: 2346-9129
ISSN-e: 2346-9129
Periodicidad: Semestral
vol. 22, e0007, 2023
Recepción: 17 Agosto 2022
Aprobación: 03 Marzo 2023
Autor de correspondencia: cdefilipis@gmail.com

Resumen: La salinización de los medios de cultivo en producciones intensivas como consecuencia del uso de agua de riego con alta conductividad eléctrica se ha incrementado en condiciones protegidas. Una estrategia para mitigar el efecto de las sales sobre los cultivos es el uso de plantas injertadas sobre un pie que tolere este estrés abiótico. El melón (Cucumis melo L.) usualmente se injerta sobre distintas especies del género Cucumis o Cucurbita, sobre híbridos interespecíficos del género Cucurbita, o sobre Lagenaria siceraria. En este estudio se comparó a través de tres experimentos el patrón de acumulación de biomasa aérea y radicular de plantines de Cucurbitamaxima Duch. x Cucurbita moschata Duch. bajo riego con solución nutritiva de diferente salinidad (0,84, 4, 6 y 8 dS.m-1). El incremento en la cantidad de sales aportadas hacia fines del ciclo evaluado determinó una menor acumulación de materia seca en las plantas con indicadores de crecimiento (RPF, TAC, ICRF e ICRP) que reflejaron esta merma, por lo que no sería recomendable el uso de este híbrido como pie de injerto para producción de plantines en condiciones salinas.
Palabras clave: pie de injerto, salinidad, melón.
Abstract: Salinization of rooting media in intensive production as a consequence of the use of irrigation water with high electrical conductivity has increased in greenhouse environments. One strategy to mitigate the effect of salts on crops is the use of plants grafted on a rootstock that tolerates this abiotic stress. Melon (Cucumis melo L.) is usually grafted using as rootstock different species of the genus Cucumis or Cucurbita, interspecific hybrids of the genus Cucurbita, or with Lagenaria siceraria. In this work, three experiments were conducted to quantify the pattern of aerial and root biomass accumulation of seedlings of Cucurbita maxima Duch. x Cucurbita moschata Duch. Four treatments were carried out using nutrient solutions of different salinity (0.84, 4.00, 6.00 and 8.00 dS m-1). The increase in the amount of salts supplied at the end of the experiments in the higher salinity treatments resulted in a lower accumulation of dry matter in the plants. Growth analysis using the leaf weight ratio (RPF), absolute growth rate (TAC), relative plant growth rate (ICRF) and leaf relative growth rate (ICRP) corroborated this decrease. The results obtained indicate that it would not be advisable to use this hybrid as a rootstock for seedling production under saline conditions.
Keywords: rootstock, salinity, melon.
Introducción
Desde el año 1941 se registran estudios para evaluar la tolerancia de los cultivos a condiciones salinas. A partir de allí, se estableció que la tolerancia genética a las sales es compleja, al estar numerosos genes involucrados en esta respuesta (Flowers, 2004).
Lo mismo sucede desde el punto de vista fisiológico. Las halófitas presentan adaptaciones morfológicas y bioquímicas que les permiten tolerar estas condiciones adversas. Hay referencias sobre algunas plantas que, si bien tienen menor potencial productivo en condiciones salino-sódicas, pueden acumular en su tallo sodio para disminuir su traslocación a las hojas, como también algunos vegetales muestran un proceso llamado “salt exclusion”, donde las células radicales, que son selectivas frente a este ión, son las responsables de la tolerancia a estas sales (Flowers, 2004; Barkla et al., 2008) o disminuir su potencial osmótico interno de tal manera que les permite continuar con la absorción de agua y sus funciones metabólicas, proceso vinculado con la síntesis de solutos orgánicos y/o acumulación de iones inorgánicos (Roy et al., 2014) mediada por la concentración de sodio en el vegetal (Zörb et al., 2014).
La concentración de sales en el medio radical puede transformarse en un problema si afecta la calidad y/o la cantidad de productos a obtener. El rendimiento disminuye si el contenido de sales supera el tolerado por las plantas al reducirse la absorción de agua por las raíces. Este efecto se acentúa si estos iones producen fitotoxicidad. Los síntomas foliares que usualmente se observan son similares a los ocasionados por una sequía (Ayers y Westcot, 1994).
Un proceso de acumulación de sales por efecto antrópico, es el caso de los suelos bajo invernadero. Concurren varios factores para que ocurra la salinización de los mismos, que involucra la calidad de agua de riego, la intensificación de la evaporación del suelo y acumulación de sales en superficie y la ausencia de lavado por precipitaciones. Entre los efectos de este proceso se encuentra la disminución del rendimiento del cultivo (El-Shraiy et al., 2011; Miralles et al., 2012; Libutti y Monteleone, 2017).
La producción de plantines mediante la siembra de las semillas sobre un sustrato en bandejas multiceldas (speedlings), se realiza utilizando volúmenes reducidos, lo que determina un incremento de la frecuencia del riego (Burés, 1997). Si el agua tiene un alto contenido de sales, se produce la salinización del sustrato, pudiendo causar efectos negativos sobre los plantines. Se encuentra ampliamente documentada la reducción en la uniformidad de la germinación y/o emergencia, una menor calidad y hasta la muerte de las plantas si la especie no es tolerante a este tipo de estrés (Niu y Cabrera, 2010;Valdés et al., 2015).
La salinización del medio de enraizamiento interfiere en la captación de agua por parte de las raíces al elevar el potencial osmótico del suelo, provocando estrés hídrico. Asimismo, se puede producir toxicidad ion especifica debido a la alta concentración de Na+ Cl- y/o el desbalance nutricional debido a los altos niveles de sodio y cloruros que reducen la absorción de K+, NO-3, PO43- causando reducción en el crecimiento y en el desarrollo de las plantas (Lams Piedra y Gonzales Cepero, 2013).
En los últimos años en Argentina se ha incrementado la producción de cucurbitáceas provenientes de plantines injertados. Estos se obtienen a partir de un “pie” y la copa o “franco” objeto de producción, ambos de la misma familia taxonómica (Lee et al., 2010; Karaaḡaḉ y Balkaya, 2013).
En el caso del melón (Cucumis melo L.), diversos autores estudiaron la importancia de la rusticidad requerida en el pie de injerto para incrementar las resistencias y/o tolerancias a diferentes factores bióticos o abióticos (SEMARNAT, 2008; Zhao et al., 2011; Schwarz et al., 2010). También se destaca el incremento en la eficiencia en el uso del nitrógeno (Colla et al., 2010) y relación de la densidad de raíces en el incremento de la absorción de agua y nutrientes (Naranjo Morán, 2014), entre otras.
El uso de melones injertados bajo condiciones salinas se justificaría considerando los efectos negativos del uso de agua de riego de baja calidad (Colla et al., 2006; Rouphael et al., 2010; Petropoulus et al., 2011; San Bautista et al., 2011; Babaj et al., 2012)
El melón puede ser injertado sobre distintas especies de Cucurbitáceas del género Cucumis o Cucurbita,sobre híbridos interespecíficos del género Cucurbita, o sobre Lagenaria siceraria. La elección de uno u otro se basa fundamentalmente en la compatibilidad del pie de injerto con el patrón o franco. A partir de esto se evalúa el vigor, su resistencia o tolerancia a estrés biótico y estrés abiótico, y que no alteren cualitativamente el producto final (King et al., 2010). El híbrido interespecífico Cucúrbita maxima Duch. X Cucúrbita moschata Duch. se adapta para sandía y melón, enfrentando correctamente situaciones productivas desfavorables. Este híbrido presenta la mayor tolerancia a la salinidad, al compararlo con otras cucurbitáceas (El-Shraiy et al., 2011). Sin embargo, existen escasos antecedentes sobre la caracterización del crecimiento del zapallo tetsukabuto (híbrido interespecífico Cucurbita maxima Duch. x Cucurbita moschata Duch.) en etapa vegetativa temprana bajo condiciones salinas tanto para condiciones óptimas de cultivo o con diferentes tipos de estrés (Santos et al., 2018).
Este trabajo tuvo como objetivo evaluar el impacto de agua de riego con diferentes concentraciones salinas del sobre el crecimiento de plantines de zapallo tetsukabuto para su potencial uso como pie de injerto.
Materiales y métodos
El material vegetal que se utilizó fue zapallo Tetsukabuto (Cucurbita maximaDuch. x Cucurbita moschata Duch.) híbrido F1 del semillero Tokita (Japón). En todos los casos el poder germinativo fue medido en el laboratorio de Fisiología Vegetal de la Universidad Nacional de Luján.
La siembra manual se hizo sobre un sustrato comercial elaborado por la empresa Terrafertil, Grow Mix MultiPro en bandejas multiceldas (speeldings) de 72 alvéolos, con un volumen de 50 cm3 cada una. Una vez realizada la siembra, las bandejas se depositaron en una mesada dentro de un invernáculo del Área Experimental de Riego y Drenaje en el Campo Experimental de la Universidad Nacional de Luján. Dado el alto poder germinativo del material vegetal (90%), se sembró una semilla por celda.
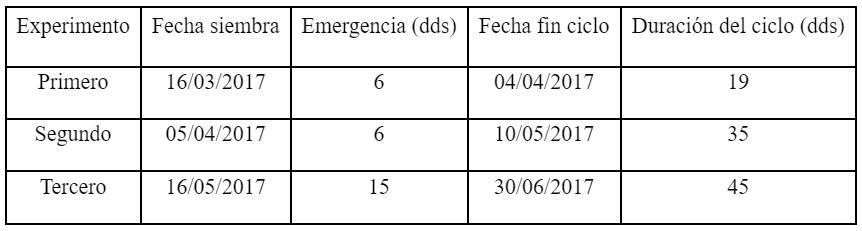
Se realizaron tres experimentos consecutivos en invernáculo con igual tecnología de cultivo (Tabla 1). Cada experimento estuvo constituido por cuatro tratamientos, determinados a partir de la conductividad eléctrica (CE) del agua de riego obtenida en todos los casos del bombeo del acuífero Arenas Puelches (CE de 0,84 dS m-1). Los tratamientos fueron: Control (agua sin adición de sales); Baja (agua con adición de cloruro de sodio, NaCl, CE 4 dS m-1); Media (agua con adición de NaCl, CE 6 dS m-1) y Alta (agua con adición de NaCl, CE 8 dS m-1). En todos los tratamientos, la CE fue determinada utilizando un medidor multiparamétrico marca Hanna modelo HI9811-5. El diseño experimental utilizado fue en bloques completamente aleatorizados. Las bandejas se dispusieron en tres bloques, estando representados todos los tratamientos en cada uno de ellos.
Las sales residuales que quedaron adsorbidas al sustrato se estimaron mediante la medición de la CE del mismo. Para ello se tomaron al azar dos muestras por repetición de cada tratamiento cuando las plantas presentaron sus cotiledones expandidos (estadio 009, código BBCH) (Meier, 2001), y luego con una frecuencia de 3 días hasta el estado de primera hoja verdadera (estadio 101, código BBCH) (Meier, 2001). En este estadio fenológico se considera que la planta se encuentra apta para su uso como pie de injerto (Lee, 1994). Las muestras de sustrato se diluyeron con agua destilada en una proporción de 5:1 y agitadas con espátula durante cinco minutos. Luego se dejaron en reposo entre 15 y 20 minutos (Karlanian, 2010). La CE se midió en el sobrenadante con un conductímetro de mesa OAKTON CON 510 Series.
De las mismas celdas utilizadas para el muestreo de sustrato, se extrajeron los plantines, a los que se les determino peso de masa seca aérea y de raíz, obtenido al secarlas en estufa a 60°C hasta peso constante, verificado con balanza analítica OHAUS EXPLORER E12140. Previo al secado, se cuantificó el diámetro del pecíolo de la primera hoja verdadera de las plantas en estadio 101, código BBCH (Meier, 2001), con un calibre digital, con un error de medición ± 0,001.
Para el análisis de crecimiento se consideraron la razón de peso foliar, RPF (Geraud y Chirinos, 1995), la tasa absoluta de crecimiento, TAC (Medina, 1977), el índice de crecimiento relativo de la planta, ICRP y el índice de crecimiento relativo foliar, ICRF (Geraud y Chirinos, 1995). A los fines de comparar los resultados de los tres experimentos se contabilizaron dos estados: de emergencia, verificada cuando la misma se observó en el 50% del total de las celdas sembradas, a la expansión de los cotiledones y de este último hasta el desarrollo de la primera hoja verdadera.
La temperatura del aire fue medida durante en todos los experimentos, registrándose la misma con paso de 30 minutos, mediante una estación meteorológica automática DAVIS modelo VANTAGE PRO 2.
Para el análisis estadístico de los datos se utilizó un diseño de parcelas divididas, en el cual se consideró a la parcela como cada ensayo y a las subparcelas como cada tratamiento (Control, Baja, Media y Alta). Posteriormente se realizó un ANAVA con el software estadístico InfoStat (Di Rienzo et al., 2020), test de Tukey para p < 0,05.
Resultados y discusión
La duración de cada uno de los experimentos fue de 19, 35 y 45 días. En el primero, la menor duración del ciclo de los plantines de Cucurbita maxima Duch. x Cucurbita moschata Duch. se originó debido a las temperaturas ocurridas, ya que los valores máximos y mínimos medios fueron de 29,9 y 15,2°C respectivamente. Estas condiciones favorecieron una mayor tasa de crecimiento de las plántulas (Maynard, 2007). Los rangos óptimos de temperaturas para las cucurbitáceas son de 21°C a 24°C durante el día y de 12°C a 18°C en la noche (FAO, 2017). Para el segundo y tercer experimento el rango de las temperaturas máximas y mínimas medias fue de 30,3-11,3°C; y 21,5-7,0C° respectivamente.
Al comparar en todos los experimentos el peso de masa seca aérea en la primera etapa fenológica (estadio 009, código BBCH, Meier, 2001) se observaron diferencias estadísticas no significativas entre los tratamientos (Tabla 2).
Posteriormente cuando se comparó el siguiente estado, comprendido entre la expansión de los cotiledones y el desarrollo de la primera hoja verdadera (estadio 101, código BBCH, Meier, 2001), se verificaron diferencias estadísticas entre todos los tratamientos (Tabla 2), correspondiendo los valores más altos de biomasa aérea al tratamiento Control en todos los experimentos. Estas diferencias fueron significativas (p<0,05) entre los tratamientos en el segundo ensayo.
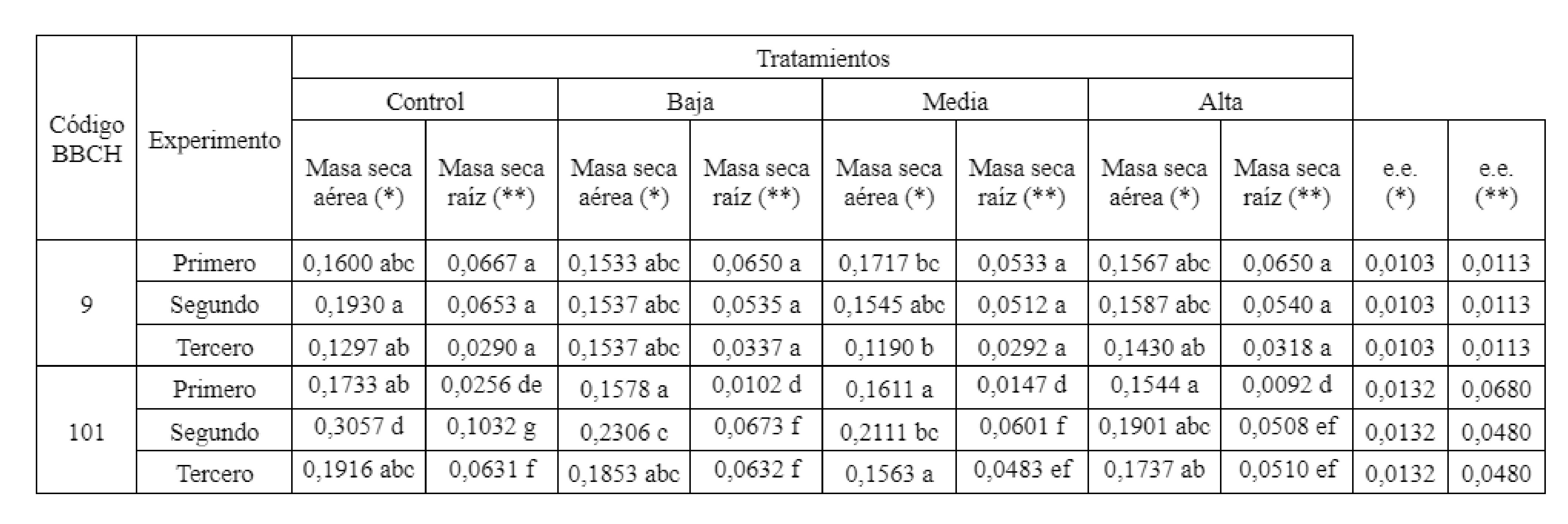
Estos resultados se corresponden con los obtenidos por Taffouo et al., (2008), quienes, trabajando con tres especies de cucurbitáceas, determinaron que Cucurbitamoschata produjo una menor cantidad de materia seca aérea y radicular al exponerla a un medio salino. El efecto supresor de la sal sobre el crecimiento de la parte aérea está dado por la alta concentración de NaCl que inhibiría el alargamiento o la división celular (Svengor et al., 2011; Munns y Tester, 2008; Greenway y Munns, 1980). Estos resultados no se observaron de manera acentuada en el primer experimento, posiblemente por la influencia de la temperatura que al incidir en un desarrollo precoz de los plantines de Cucurbita maxima (Duch). x Cucurbita moschata (Duch.). Esta situación puede haber favorecido un menor tiempo de exposición a las condiciones salinas reduciéndose así el efecto de este estrés abiótico, y disminuyendo la concentración de Na. y Cl. en el interior de la planta (Modarelli et al., 2020).
Durante el estadio 009 BBCH (Meier, 2001), los pesos de masa seca radicular presentaron no diferencias significativas (p<0,05) entre los tratamientos regados con solución salina para los últimos dos experimentos realizados (Tabla 2). Al analizar la etapa mediada entre expansión de los cotiledones y el desarrollo de la primera hoja verdadera, se observó que la dinámica de acumulación de materia seca en raíz fue similar al de la parte aérea, con diferencias significativas entre el tratamiento Control del segundo experimento y los otros tratamientos (Tabla 2).
Los tratamientos utilizados provocaron un incremento de la concentración salina del medio de cultivo, estimada a través de la conductividad eléctrica (CE), registrando al final del ciclo evaluado una media de los experimentos de 0,62, 2,57 3,86, 5,46 dS m-1 (± 0,46) para los tratamientos Control, Baja, Media y Alta. Esto incidió negativamente en la traslocación de asimilados a los órganos del plantín con el avance del ciclo evaluado al mostrar menor materia seca en el estadio 0101 BBCH (Meier, 2001) los tratamientos regados con solución nutritiva de CE modificada por el agregado de NaCl (Tabla 2). Edelstein et al., (2010), registraron en plantas de Cucurbita maxima (Duch). x Cucurbita moschata (Duch.) a los 30 días posteriores al trasplante, una mayor acumulación de materia seca en la parte aérea respecto a la radicular, cuando se regaron con una solución salina de 4,5 dS m-1, tendencia similar a la obtenida en estos ensayos.
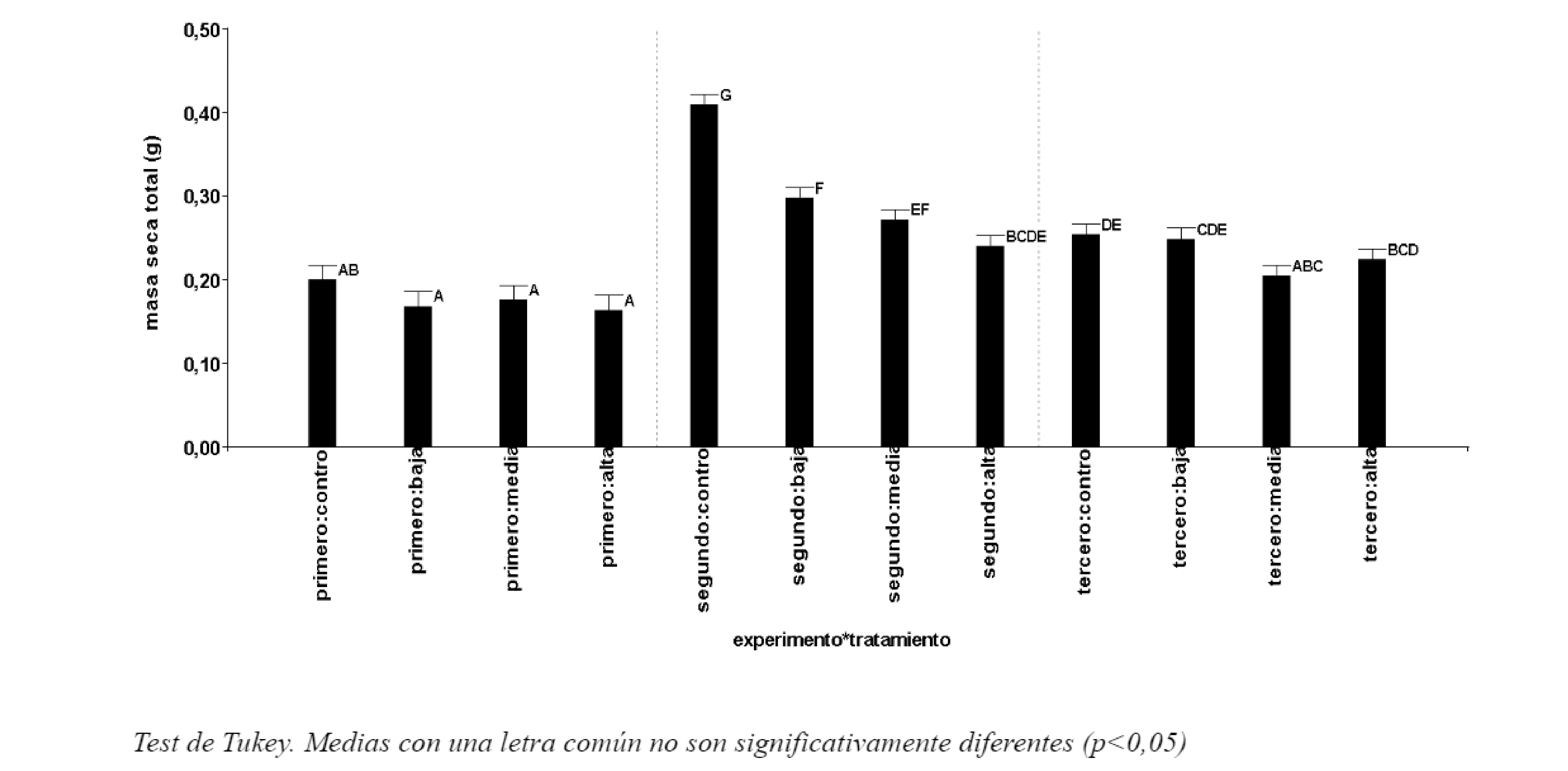
Al finalizar el experimento en el estado 101 código BBCH (Meier, 2001) las mediciones del peso de masa seca total permitieron observar la influencia negativa de las sales sobre el crecimiento de los plantines (Figura 1). Esto pudo determinarse a través de los resultados obtenidos de la CE del medio en todos los experimentos. Además, se pudo observar el efecto favorable del rango térmico durante el segundo experimento. Las diferencias significativas observadas siguieron la misma tendencia que los pesos parciales de las estructuras vegetales antes evaluadas.
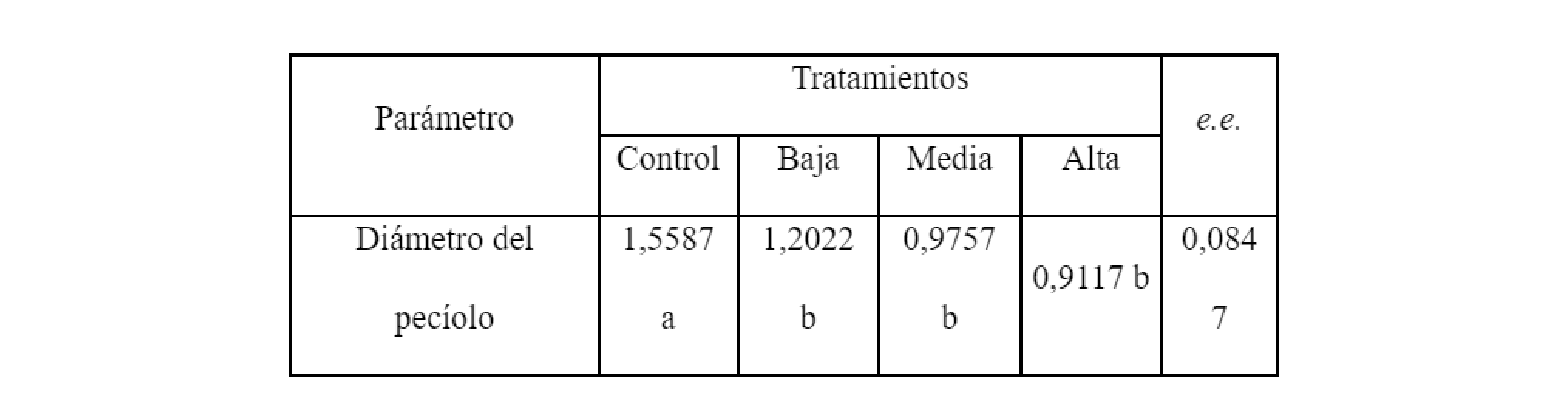
La operación de injerto en especies hortícolas, cuando la técnica es de aproximación, requiere que los diámetros de los talluelos sean similares en cuanto a su diámetro. Esta variable, evaluada como media de los tres experimentos, mostró diferencias significativas entre el Control y los tratamientos al ser afectada levemente por el incremento de sales en la solución de riego (Tabla 3) posiblemente por una menor acumulación de materia seca.
A partir de los valores de masa seca obtenidos en los experimentos se cuantificó el crecimiento de los plantines, a través de tasas e índices que no mostraron diferencias significativas entre los tratamientos salinos ensayados sobre los plantines de Cucurbita maxima Duch. x Cucurbitamoschata Duch. (Tabla 4).
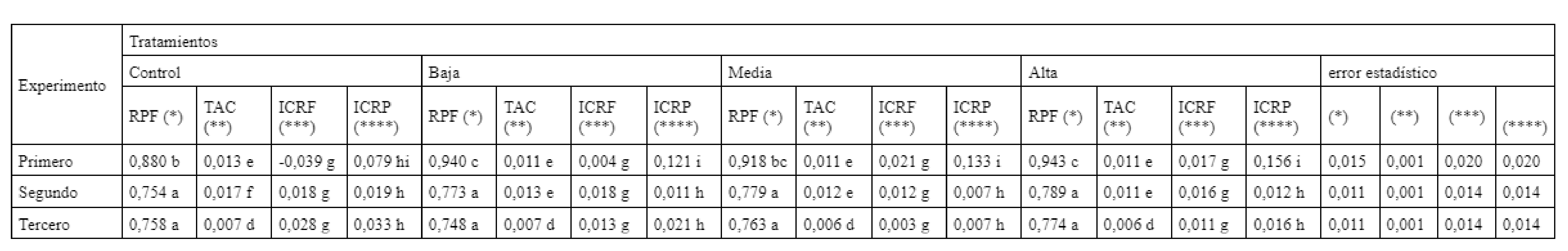
RPF: razón de peso foliar
TAC: tasa absoluta de crecimiento
ICRF: índice de crecimiento relativo foliar
ICRP: índice de crecimiento relativo de la planta
Resulta destacable que en las variables de crecimiento tasa absoluta de crecimiento (TAC) e índice de crecimiento relativo de la planta (ICRP) (Tabla 4), al aumentar el contenido salino en el agua de riego, estos índices decrecen, posiblemente como consecuencia del incremento en la acumulación de sales en el sustrato. Esto se hace más notorio en el ICRP al expresar el incremento en masa seca de la planta en un intervalo de tiempo dado, tomando como referencia el valor inicial de la masa seca producida y acumulada (Tabla 4). El ICRF, que cuantifica la evolución de la masa seca aérea en función de la masa seca total, decrece cuando el riego se realiza con solución de CE superior a los 4 dS m-1. La media de la relación entre la biomasa aérea y la biomasa total (RPF) de los plantines fue similar para todos tratamientos salinos, con valores superiores al tratamiento Control (Tabla 4), dado que la condición salina impuesta impactó en mayor medida sobre la masa seca radicular que sobre la aérea (Tabla 2).
Estos resultados harían suponer que en Cucurbita maxima Duch. x Cucurbita moschata Duch. no actuaría adecuadamente el mecanismo de exclusión de Cl- y Na+ (Edelstein and Ben-Hur, 2012), siendo este último uno de los principales atributos para un adecuado pie de injerto en pepino y melón (Huang et al., 2010) y que hubiese mitigado el efecto negativo de las sales de NaCl en el crecimiento (Modarelli et al., 2020; Munns y Tester, 2008).
En cucurbitáceas se observó que la alta concentración de sodio en el agua de riego produjo un gradual incremento en el tenor salino del medio radical, respondiendo la planta a esta modificación con una reducción de su área foliar y del crecimiento del tallo (Sivritepe et al., 2005), del potencial agua de las hojas, de la conductancia estomática (Kusvuran, 2012) y de variables morfogenéticas tales como la razón de área foliar y el área foliar específica. La reducción en la producción de materia seca se visualiza en una menor tasa de crecimiento relativo. Generalmente, este detrimento se puede deber a las altas concentraciones salinas en el apoplasto, provocando la deshidratación de las células y tejidos. Cómo se mencionó anteriormente, esta tendencia decreciente es la que se observó para el ICRP, índice que cuantifica la tasa de crecimiento relativo medido en los plantines de Cucurbita maxima Duch. x Cucurbita moschata Duch. en los tres experimentos.
Estos resultados preliminares deberán complementarse con estudios que permitan determinar la morfología del sistema radical. Rasgos morfológicos y arquitectónicos de la raíz, como el diámetro, la densidad y distribución, que afectan la absorción de agua (Ho et al., 2005; Comas et al., 2013; Mickelbart et al., 2015) y tolerancia a la salinidad (Bertucci et al., 2018). Hasta la fecha, se han realizado pocas investigaciones para comparar los sistemas de raíces de portainjertos de melón, por lo que sería de interés comparar atributos morfológicos y fisiológicos que permitan explicar las diferencias entre C. maxima Duch. x C. moschata Duch. y otros pies de injertos utilizados como C. ficifolia y C. moschata (Chen and Wang, 2008) y Lagenaria siceraria (Yetisir and Uygur, 2010).
Conclusiones
Al finalizar los experimentos, el incremento de la salinidad redujo la tasa de incremento de materia seca en las plántulas de Cucurbita maxima Duch. x Cucurbita moschata Duch.
En fase que media entre la expansión de los cotiledones y el desarrollo de la primera hoja verdadera, la salinidad provocó un menor acúmulo de materia seca en comparación con el tratamiento control.
El diámetro del epicótile resultó inferior en los tratamientos de salinidad en comparación con el tratamiento control.
Los resultados obtenidos indicarían que Cucurbita maxima Duch. x Cucurbita moschata Duch. no es un genotipo tolerante a salinidad en etapa vegetativa temprana, no siendo indicado para utilizar en producciones con destino a pie de injerto en presencia de este estrés abiótico.
Agradecimientos
A la Srta. María Eugenia Elli, técnica química del laboratorio del Campo Experimental de la Universidad Nacional de Luján. El presente trabajo contó con financiamiento del Departamento de Tecnología de la Universidad Nacional de Luján.
Referencias bibliográficas
Ayers, R.S.; Westcot, D.W. (1994). Water quality for agricultura. FAO Irrigation and Drainage paper Nº 29. https://www.fao.org/3/t0234e/t0234e.pdf.
Babaj, I.; Sallaku, A.; Balliu, A. (2012). The effect of plug size and grafting method on the dry matter partitioning and stand establishment rate of grafted cucumber seedlings under saline conditions. Acta Horticulturae 960, 225-230. https://doi.org/10.17660/ActaHortic.2012.960.32
Barkla, B; Vera-Estrella, R.; Balderas, E.; Pantoja, O. (2008) Mecanismos de tolerancia a la salinidad en plantas En: F. Rebolledo y A. López, (eds.). Una ventana al quehacer científico (pp 263-272). Instituto de Biotecnología, UNAM. México, D.F.
Bertucci, M.B., Suchoff, D.H., Jennings, K.M., Monks, D.W., Gunter, C.C., Schultheis, J.R., Louws, F.J. (2018). Comparison of root system morphology of Cucurbit rootstocks for use in Watermelon grafting. HortTechnology 28(5),629-636.
Burés, S. (1997). Sustratos. Ediciones Agrotécnicas S.L. Madrid, España.
Chen, G., Wang, R. (2008). Effects of salinity on growth and concentration of sodium, potassium and calcium in grafted cucumber seedling. Acta Horticulturae 771,217-224.
Colla, G.; Rouphael, Y.; Cardarelli, M.; Massa, D.; Salerno, A. (2006). Yield, fruit quality and mineral composition of grafted melon plants grown under saline conditions. Journal of Horticultural Science & Biotechnology 81(1), 146-152. https://doi.org/10.1080/14620316.2006.11512041
Colla, G.; Rouphael, Y.; Cardarelli, M.; Massa, D.; Salerno, A. (2010). Role of grafting in vegetable crops grown under saline conditions. Scientia Horticulturae 127(2), 147-155. https://doi.org/10.1016/j.scienta.2010.08.004
Comas, L., Becker, S., Cruz, V., Byrne, P., Dierig, D. (2013). Root traits contributing to plant productivity under drought Frontiers in Plant Science. 4(442),1-16.
Di Rienzo J., Casanoves F., Balzarini M., Gonzalez L., Tablada M., Robledo C. (2020) InfoStat versión 2020. Centro de Transferencia InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. URL http://www.infostat.com.ar
Edelstein, M.; Plaut, Z.; Ben-Hur, M. (2010). Sodium and chloride exclusion and retention by non-grafted and grafted melon and Cucurbita plants. Journal of Experimental Botany 62(1),177-184 https://doi.org/10.1093/jxb/erq255
Edelstein, M., Ben-Hur, M. (2012). Use of grafting to mitigate chemical stresses in vegetables under arid and semiarid conditions. Advances in Environmental Research 20,163-179.
El-Shraiy, A.; Mostafa, M.; Zaghlool, S.; Shehata., S. (2011). Physiological Aspect of NaCl-salt Stress Tolerant among Cucurbitaceous Cultivars. Australian Journal of Basic and Applied Sciences 5(11), 62-71. http://www.ajbasweb.com/old/ajbas/2011/November-2011/62-71.pdf
Flowers, T. (2004). Improving crop salt tolerance. Journal of Experimental Botany 55(396), 307-319. https://doi.org/10.1093/jxb/erh003
Geraud, F.; Chirinos, D. (1995). Desarrollo de la planta de tomate Lycopersicon esculentum cv. Rio Grande en la zona de Rio Limón del Estado Zulia, Venezuela. II. Índice de crecimiento relativo, razón de peso foliar y gamma. Revista Facultad de Agronomía 12, 15-23.
Greenway, H.; Munns, R. (1980) Mechanisms of salt tolerance in nonhalophytes. Annual Review of Plant Physiology 31(1), 149-190. https://doi.org/10.1146/annurev.pp.31.060180.001053
Ho, M., Rosas, J., Brown, K., Lynch, J. (2005). Root architectural tradeoffs for water and phosphorus acquisition Funct. Plant Biol. 32, 737-748.
Huang, Y., Bie, Z., He, S., Hua, B., Zhen, A., Liu, Z. (2010). Improving cucumber tolerance to major nutrients induced salinity by grafting onto Cucurbita ficifolia. Environmental and Experimental Botany 69, 32-38.
Karaaḡaḉ, O.; Balcaya, A. (2013). Interspecific hybridization and hybrid seed yield of winter squash (Cucurbita maxima Duch.) and pumpkin (Cucurbita moschata Duch.) lines for rootstock breeding. Scientia Horticulturae 149(4), 9 - 12. https://doi.org/10.1016/j.scienta.2012.10.021
Karlanian, M. (2010). Como medir pH y conductividad eléctrica. Economia & Viveros. https://www.economiayviveros.com.ar/archivo/enero2010/como_hacerlo.html
King, S.; Davis, A.; Zhang, X.; Crosbya. K. (2010). Genetics, breeding and selection of rootstocks for Solanaceae and Cucurbitaceae. Scientia Horticulturae 127(2), 106-111. https://doi.org/10.1016/j.scienta.2010.08.001
Kusvuran, S. (2012). Effects of drought and salt stresses on growth, stomatal conductance, leaf water and osmotic potentials of melon genotypes (Cucumis melo L.). African Journal of Agricultural Research 7(5), 775-781. https://doi.org/10.5897/AJAR11.1783
Lams Piedra, A.; Gonzales Cepero, M. (2013). La salinidad como problema en la agricultura: la mejora vegetal una solución inmediata. Cultivos Tropicales 34(4),31-42. http://scielo.sld.cu/pdf/ctr/v34n4/ctr05413.pdf
Lee, J. (1994). Cultivation of grafted vegetables I. Current status, grafting methods, and benefits. HortScience 29(4),235-239. https://doi.org/10.21273/HORTSCI.29.4.235
Lee, J.; Kubota, C.; Tsao, S.; Bie, Z.; Hoyos Echevarría, P.; Morra, L.; Oda, M. (2010). Current status of vegetable grafting: Diffusion, grafting techniques, automation. Scientia Horticulturae 127 (2), 93-105. https://doi.org/10.1016/j.scienta.2010.08.003
Libutti, A.; Monteleone, M. (2017). Soil vs. groundwater: The quality dilemma. Managing nitrogen leaching and salinity control under irrigated agriculture in Mediterranean conditions. Agricultural Water Management 186,40-50. https://doi.org/10.1016/j.agwat.2017.02.019
Maynard, L. (2007). Cucurbit crop growth and development. En: University of Purdue Conference Proceedings, Indiana, United States of America. https://www.agry.purdue.edu/cca/2007/2007/Proceedings/Liz%20Maynard-CCA%20proceedings%201_KLS.pdf
Medina, E. (1977). Distribución de asimilados y análisis de crecimiento. En: Instituto Venezolano de Investigaciones Cientificas. Caracas, Venezuela. .Introducción a la ecofisiología vegetal (pp. 51-56).
Meier, U. (2001). Estadios de las plantas mono y dicotiledóneas [BBCH Monografía]. Centro Federal de Investigaciones Biológicas para Agricultura y Silvicultura. https://www.julius-kuehn.de/media/Veroeffentlichungen/bbch%20epaper%20span/page.pdf
Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. (2015). Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nature Reviews Genetic 16(4) ,237-251.
Miralles J.; Valdes, R.; Martínez-Sánchez, J.; Bañon, S. (2012). Pot-in-pot reduces salinity, chloride uptake, and maintains aesthetic value in Euonymus japonicas Thunb. under saline irrigation. HortScience 47(5), 607-613. https://doi.org/10.21273/HORTSCI.47.5.607
Modarelli, G.; Rouphael, Y.; De Pascale, S.; Öztekin, G.; Tüzel, Y.; Orsini, F.; Gianquinto, G. (2020) Appraisal of salt tolerance under greenhouse conditions of a Cucurbitaceae Genetic Repository of potential rootstocks and scions. Agronomy 10(7), 967. https://doi.org/10.3390/agronomy10070967
Munns, R.; Tester, M. (2008). Mechanisms of salinity tolerance. Annual Review of Plant Biology 59(1), 651-681. https://doi.org/10.1146/annurev.arplant.59.032607.092911
Naranjo Morán, J. (2014). Evaluación de la tolerancia a la salinidad de cucurbitáceas silvestres del Ecuador y sus potenciales usos como patrones en injertos de cucurbitáceas comerciales. (Tesis de grado, Facultad de Ingeniería en Mecánica y Ciencias de la Producción FIMCP – ESPOL, Guayaquil. Ecuador). http://www.dspace.espol.edu.ec/handle/123456789/25205
Niu, G.; Cabrera, R. (2010). Growth and physiological responses of landscape plants to saline water irrigation: A review. HortScience 45(11), 1605-1609 https://doi.org/10.21273/HORTSCI.45.11.1605
Petropoulos, S.; Olympios, C.; Ntatsi, G.; Akoumiankis, C.; Passam, H. (2011). Effects of grafting and salinity on agronomic characteristics and ion uptake by cucumber plants grown in a closed hydroponic system. En: International Symposium on Advanced Technologies and Management Towards Sustainable Greenhouse Ecosystems: Greensys2011 952 (pp. 637-643). https://doi.org/10.17660/ActaHortic.2012.952.80
Rouphael, Y.; Schwarz, D.; Krumbein, A.; Colla, G. (2010). Impact of grafting on product quality of fruit vegetables. Scientia Horticulturae 127(2), 172-179. https://doi.org/10.1016/j.scienta.2010.09.001
Roy, S.; Negrao, S.; Tester, M. (2014). Salt resistant crop plants. Current Opinion in Biotechnology 26, 115-124. https://doi.org/10.1016/j.copbio.2013.12.004
San Bautista, A.; Calatayud, A.; Nebauer, S.; Pascual, B.; Maroto, J.; López-Galarza, S. (2011). Effects of simple and doublé grafting melón plants on mineral absorption, photosynthesis, biomass and yield. Scientia Horticulturae 130(3), 575-580. https://doi.org/10.1016/j.scienta.2011.08.009
Santos, A.; Sá, F.; Souto, L.; Silva, M.; Moreira, R.; Lima, G., Mesquita, E. (2018). Tolerance of varieties and hybrid of pumpkin and squash to salt stress. Journal of Agricultural Science 10(1), 38-44. https://doi.org/10.5539/jas.v10n1p38
Schwarz, D.; Rouphael, Y.; Colla, G.; Venema, J. (2010). Grafting as a tool to improve tolerance of vegetables to abiotic stresses: Thermal stress, wáter stress and organic pollutants. Scientia Horticulturae 127(2), 162-171. https://doi.org/10.1016/j.scienta.2010.09.016
SEMARNAT (Secretaría de Medio Ambiente y Recursos Naturales). (2008). El injerto en el cultivo de melón y sandía como alternativa al uso de bromuro de metilo. http://app1.semarnat.gob.mx:8080/sissao/images/pdf/MELONYSANDIA-CO.pdf
Sivritepe, H. O.; Sivritepe,N.; Eriş, A.; Turhan, E. (2005). The effects of NaCl pre-treatments on salt tolerance of melons grown under long-term salinity. Scientia Horticulturae 106,568-581. https://doi.org/10.1016/j.scienta.2005.05.011
Svengor, S.; Yasar, F.; Kusvuran, S.; Ellialtoglu, S. (2011). The effect of salt stress on growth, clorophyll content, lipid peroxidation and antioxidative enzymes of pumpkin seedling. African Journal of Agricultural Reaserch 6(21), 4920-4924. https://doi.org/10.5897/AJAR11.668
Taffouo, V.; Djiotie, N.; Kenné, M.; Din, N.; Priso, J.; Dibong, S.; Akoa, A. (2008). Effects of salt stress on physiological and agronomic characteristics of three tropical cucurbit species. Journal of Applied Biosciences 10,434-441. http://www.m.elewa.org/JABS/2008/10(1)/2.pdf
Valdés R.; Franco, J.; Sánchez Blanco, M.; Bañón, S. (2015). Relationships among electrical conductivity measurements during saline irrigation of potted Osteospermum and their effects on plant growth. The Journal of Horticultural Science and Biotechnology 90(5), 571-577. https://doi.org/10.1080/14620316.2015.11668716
Zhao, X.; Guo, Y.; Huber, D.; Lee, J. (2011). Grafting effects on postharvest ripening and quality of 1-methylcyclopropene-treated muskmelon fruit. Scientia Horticulturae 130(3), 581-587. https://doi.org/10.1016/j.scienta.2011.08.010
Yetisir, H., Uygur, V. (2010). Response of grafted watermelon onto different gourd species to salinity stress. Journal of Plant Nutrition 33,315-327.
Notas de autor
cdefilipis@gmail.com

