

Efecto de ácido naftalenacético, ácido indol-3-butírico y ácido indol-3-acético en la respuesta rizogénica in vitro de Neltuma alba
Effect of naphthaleneacetic acid, indole-3-butyric acid, and indole-3-acetic acid on the in vitro rhizogenic response of Neltuma alba
 Edgardo José Carloni carloni.edgardo@inta.gob.ar
Karina Grunberg grunberg.karina@inta.gob.ar
Edgardo José Carloni carloni.edgardo@inta.gob.ar
Karina Grunberg grunberg.karina@inta.gob.ar
Revista FAVE Sección Ciencias Agrarias
Universidad Nacional del Litoral, Argentina
ISSN: 2346-9129
ISSN-e: 2346-9129
Periodicidad: Semestral
núm. 23, e0026, 2024
Recepción: 10 octubre 2023
Aprobación: 15 febrero 2024

Resumen: Neltuma alba, conocido como “algarrobo blanco”, es una especie nativa importante en la ecorregión región del Chaco y especialmente en Argentina debido a su valor biológico, productivo y cultural. La sobreexplotación maderera está generando una pérdida de la diversidad genética de la especie. La propagación vegetativa in vitro se presenta como una herramienta biotecnológica para la conservación de los recursos genéticos forestales. El objetivo de este estudio fue evaluar los efectos de diferentes tipos y concentraciones de auxinas en la respuesta rizogénica in vitro de N. alba. Los resultados indican que los explantes tienen capacidad para desarrollar raíces adventicias con o sin la presencia de fitorreguladores, aunque concentraciones mayores de auxinas, inducen una mejor respuesta rizogénica. El subcultivo de los explantes en un medio sin auxinas, después de 7 días de exposición a 4 mg/l de IBA, ha demostrado reducir la formación de callos y minimizar las malformaciones en las raíces. Esta metodología de trabajo permite alcanzar una tasa de sobrevivencia del 75% de las plantas ex vitro.
Palabras clave: auxinas, Neltuma sp. conservación, raíces adventicias, explantes.
Abstract: Neltuma alba, known as “algarrobo blanco” is an important native species in the Chaco ecoregion, particularly in Argentina, due to its biological, productive, and cultural value. Overexploitation of its timber is causing a loss in the species' genetic diversity. Vegetative propagation in vitro is emerging as a biotechnological tool for conserving forest genetic resources. This study aimed to assess the effects of different types and concentrations of auxins on the in vitro rooting response of N. alba. The results indicate that explants have the ability to develop adventitious roots with or without the presence of plant growth regulators, though higher concentrations of auxins induce a better rooting response. Subculturing explants in an auxin-free medium after 7 days of exposure to 4 mg/l of IBA has shown to reduce callus formation and minimize root malformations. This working methodology allows achieving a 75% survival rate of ex vitro plants.
Keywords: auxins, Neltuma sp., conservation, adventitious roots, explants.
INTRODUCCIÓN
Neltuma alba (Griseb.) (ex género Prosopis), conocido comúnmente como "algarrobo blanco", es una de las especies forestales nativas con mayor distribución geográfica dentro de la ecorregión del Chaco (Hughes et al., 2022; Vega et al., 2021). En Argentina, se encuentra en las provincias de Chaco, Córdoba, Corrientes, Entre Ríos, Formosa, Jujuy, Salta, Santa Fe, Santiago del Estero y Tucumán. Es considerado un árbol multipropósito debido a su alto valor biológico, productivo y cultural (Verga y López Lauenstein, 2021). En el país, la actividad forestal asociada a esta especie se centra en la extracción de madera de los bosques nativos (Venier et al., 2013). La sobreexplotación maderera y la tala selectiva de los árboles de elite está generando una pérdida de diversidad genética en las poblaciones naturales de la especie (Verga y López Lauenstein, 2021). En la actualidad, los árboles de élite son difíciles de encontrar, situándose en general en áreas aisladas, lo que subraya la necesidad de desarrollar herramientas biotecnológicas para conservar los recursos genéticos forestales.
La técnica de propagación vegetativa a través del cultivo in vitro de tejidos vegetales se presenta como una herramienta para la conservación de los recursos genéticos forestales (Morales-Domínguez et al., 2019). Diversos programas de mejora genética han integrado el cultivo in vitro con el objetivo de clonar árboles adultos de poblaciones naturales, conservarlos y propagarlos vegetativamente en entornos controlados (Brondani et al., 2012). Estas estrategias permiten mantener y preservar la diversidad genética de especies forestales de interés, asegurando la disponibilidad de material vegetal de calidad y la continuidad de las características deseables en futuras generaciones (Hartmann et al., 2011).
En el caso particular de Neltuma sp., se han realizado diversos avances en cuanto al establecimiento in vitro utilizando diferentes tipos de explantes, como ápices de tallos, brotes axilares y segmentos uninodales (Castillo y Bovo, 2000; Jordan, 1987; Tabone et al., 1986). Los resultados más favorables se han obtenido al cultivar segmentos uninodales en medio basal MS (Murashige y Skoog, 1962), a una temperatura de 25°C y bajo un fotoperíodo de 18:6 (Luz:Oscuridad) (Jordan, 1987; Shekhawat et al., 1993). Diversas especies del género, como N. alba, N. alpataco, N. caldenia, N. chilensis, N. juliflora, N. pallida, N. limensis y N. laevigata, han sido propagadas vegetativamente a través del cultivo in vitro (Arce y Balboa, 1991; Boeri y Sharry, 2018; Jordan et al., 1985; Minchala-Patiño et al., 2014; Morales-Domínguez et al., 2019; Nandwai y Ramawat, 1991; Rivera Curi et al., 2020; Verdes, 2007). No obstante, algunos autores sugieren que en el caso específico de N. alba se requieren nuevos estudios para implementar eficazmente la técnica del cultivo in vitro. En particular, haciendo énfasis en mejorar la viabilidad de los brotes cultivados in vitro, la producción de raíces adventicias en los explantes y la exitosa transferencia al invernadero de las plántulas obtenidas in vitro (Tabone et al., 1986; Green et al., 1990).
En cuanto al proceso de formación de raíces adventicias, las auxinas son los reguladores de crecimiento que desempeñan un papel importante. Entre las más utilizadas en Neltuma sp. se encuentran el ácido indol-3-acético (AIA), el ácido naftalenacético (ANA) y el ácido indol-3-butírico (AIB) (Arce y Balboa, 1991; Minchala-Patiño et al., 2014; Morales-Domínguez et al., 2019; Nandwai y Ramawat, 1991; Rivera Curi et al., 2020; Verdes, 2007). Es conocido que la respuesta rizogénica está regulada por el tipo y la concentración de auxina presente en el medio de cultivo, y que el modo de acción y la eficacia de cada una de ellas pueden variar entre especies (Batchelor et al., 1989; Jordan et al., 1985; Goyal y Arya, 1984). Incluso, algunos autores mencionan diferencias en la respuesta rizogénica dentro de una misma especie, dependiendo del origen del material vegetal (Arce y Balboa, 1991).
El objetivo de este estudio fue evaluar los efectos de diferentes tipos y concentraciones de auxinas en la respuesta rizogénica in vitro de una población de N. alba.
MATERIALES Y MÉTODOS
Material vegetal
Se utilizaron semillas de N. alba provenientes de una muestra de una población ubicada en la localidad de Campo Durán (22°12'1"S, 63°40'33"O), en la provincia de Salta, Argentina. Estas semillas fueron suministradas por el Banco Nacional de Germoplasma de Prosopis de la Facultad de Ciencias Agropecuarias de la Universidad Nacional de Córdoba (BNGP-FCA-UNC), y posteriormente almacenadas en el Instituto de Fisiología y Recursos Genéticos Vegetales (IFRGV) del Instituto Nacional de Tecnología Agropecuaria (INTA) en Córdoba, Argentina.
Cultivo in vitro
Para la introducción al cultivo in vitro, se sometieron aproximadamente 25 semillas de N. alba a un proceso de escarificación utilizando papel de lija (Venier et al., 2015). La desinfección de las semillas se llevó a cabo mediante agitación constante, sumergiéndolas en etanol al 70 % (v/v) durante 1 minuto, seguido de una inmersión en una solución comercial de hipoclorito de sodio al 5 % (35 g/l) con dos gotas de Tween 20 durante 30 minutos. Posteriormente, se realizaron cinco enjuagues con agua destilada estéril dentro de una cámara de flujo laminar.
Las semillas desinfectadas fueron cultivadas in vitro en un medio de germinación compuesto por sacarosa al 1 % (p/v) y las sales del medio basal Murashige y Skoog (MS) a la mitad de su concentración normal (1/2x) (Shekhawat et al., 1993). Cada semilla fue transferida a un tubo de vidrio de 2 cm de diámetro x 11 cm de altura que contenía 10 ml de medio de germinación. Posteriormente, los tubos fueron colocados en la cámara de incubación en condiciones controladas de temperatura (25°C ± 2°C) y fotoperíodo (16 horas de luz a una intensidad de 95.46 umol m. s-1, seguidas de 8 horas de oscuridad).
Después de 45 días, las plántulas obtenidas fueron seccionadas en segmentos que contenían dos a tres entrenudos, con una longitud promedio de 2,5 ± 0,5 cm (Fig. 1.a). Cada segmento individual (explante) fue transferido al mismo medio de germinación, al que se le aplicaron diferentes tratamientos: tres auxinas como reguladores de crecimiento (IBA - I-5386 de SIGMA., ANA - N-0640 de SIGMA., AIA - I-2886 de SIGMA.) en siete concentraciones diferentes (0; 1; 0,5; 1; 2; 4; 6 y 8 mg/l) para cada una de ellas. El mismo medio de cultivo de germinación sin auxinas se utilizó como control. Los tubos fueron distribuidos aleatoriamente en la cámara de incubación, manteniendo condiciones controladas de temperatura (25°C ± 2°C) y fotoperíodo (16 horas de luz a una intensidad de 95,46 umol m. s-1, seguidas de 8 horas de oscuridad). Se evaluaron 19 ± 1 repeticiones para cada auxina y concentración, así como para la condición control, considerando cada tubo de cultivo con un explante como la unidad experimental. Todos los medios utilizados fueron ajustados a un pH de 5,8, se les añadió 2,5 g/l de gellan gum (Culture Gel™ Type 1 - PhytoTechnology) y se autoclavaron durante 20 minutos a 120°C.
Tras la evaluación inicial del efecto de las auxinas y sus concentraciones en la respuesta rizogénica, se llevó a cabo un nuevo ensayo utilizando el mismo medio de cultivo de germinación y con el agregado de 4 mg/l de AIB. Los explantes fueron cultivados en este medio durante siete días y posteriormente fueron subcultivados en el mismo medio de germinación durante 30 días más, esta vez sin la presencia de la auxina. Se realizaron 20 repeticiones, considerando cada tubo de cultivo con un explante como la unidad experimental. Este ensayo fue realizado manteniendo condiciones controladas de temperatura y fotoperíodo, como se mencionó anteriormente. Finalmente, las plántulas enraizadas de este ensayo fueron trasplantadas a tubos plásticos de 400 cm. (Tubete R400, Dassplastic) que contenían una mezcla de tierra y arena (1:1) como sustrato y trasladadas a un invernadero (32°C ± 2°C) cubierto con un paño de sombra de aluminio al 50 % reflectante metalizado (HDPE). Para mantener un ambiente saturado de humedad durante el período de rusticación, se colocaron frascos de vidrio (360 cm.) sobre las plántulas después del trasplante. Tras 15 días, se retiraron los frascos y, después de 30 días, se determinó el porcentaje de sobrevivencia de las plantas ex vitro.
Variables analizadas
Para determinar si el tipo de auxina y las concentraciones utilizadas en los medios tienen efecto sobre la respuesta rizogénica, fueron evaluadas, a los 30 días de iniciado el cultivo in vitro, las siguientes variables: el porcentaje de explantes con raíz, el número de raíces (NR), el peso fresco (PFR), el peso seco de raíces (PSR), el peso fresco (PFC) y peso seco de los callos (PSC) por explante. El porcentaje de explantes con raíz fue calculado considerando el número de explantes que desarrollaron al menos una raíz en relación con el total de explantes utilizados en cada tratamiento, multiplicado por 100. La determinación del NR se basó en el recuento directo de raíces en los explantes mediante el uso de una lupa binocular (Zeiss Stemi 1000). El cálculo del PFR y el PFC fue realizado extrayendo las raíces y los callos de los explantes, respectivamente, y pesándolos, utilizando una balanza analítica (Ohaus Pioneer PX). Posteriormente, tanto las raíces como los callos fueron sometidos a un proceso de secado en una estufa a 78°C hasta alcanzar un peso constante, lo que permitió la determinación del PSR y el PFC, respectivamente. El método de muestreo fue destructivo, evaluándose 19 ± 1 repeticiones para cada combinación de auxina y concentración.
Análisis estadístico
El porcentaje de explantes con raíz fue analizado mediante tablas de contingencia, utilizando sus marginales para estimar la independencia de la variable mediante la prueba X2. El efecto de las auxinas y concentraciones se consideraron efectos fijos. Para evaluar la variable NR, se utilizaron modelos lineales generalizados mixtos (MLGM) con una distribución Poisson y una función de enlace log. En relación a las variables PFR, PSR, PFC y PSC, se utilizó un modelo lineal general (MLG), y se realizó una corrección para la homogeneidad y normalidad mediante el uso de varIden. El análisis estadístico para las variables NR, PFR, PSR, PFC y PSC fue llevado a cabo utilizando un modelo con interacción entre las diferentes fuentes de variación (auxinas*concentraciones), y las diferencias entre medias fueron evaluadas mediante el test DGC (Di Rienzo et al., 2002). Todos estos análisis fueron realizados utilizando el software estadístico InfoStat (Di Rienzo et al., 2012).
RESULTADOS
En cuanto al proceso de formación de raíces (Figura 1), a partir del tercer día después de la introducción del explante en el tubo de cultivo, se observó una hinchazón en la parte basal del explante. Alrededor del quinto día, se formó una rajadura en la epidermis del tallo a través de la cual emergió el ápice radicular (Figura 1.b). Aproximadamente al séptimo día, las raíces adventicias se volvieron visibles a simple vista (Figura 1.c y d). Posteriormente, emergieron otras raíces adventicias del explante, dando lugar a una etapa de elongación (Figura 1.e). Pasados aproximadamente 20 días, fue posible observar el número final de raíces adventicias formadas (NR) (Figura 1.f).

Los resultados mostraron que los explantes jóvenes tenían la capacidad de regenerar raíces adventicias tanto en presencia como en ausencia de auxinas en el medio (Tabla 1). Al emplear las concentraciones como criterio de clasificación y al particionar por auxinas, se observó un incremento en el porcentaje de explantes con raíces a medida que las concentraciones aumentaban, tanto con AIA (P < 0,0001) como con ANA (P < 0,0001) (Tabla 1). Respecto a esta variable, los valores de X2 no revelaron diferencia significativa en el AIB (P = 0,4143). Por otro lado, al utilizar las auxinas como criterio de clasificación y particionar por concentraciones, los valores de X2 indicaron que, en general, concentraciones ≥ a 4 mg/l (a excepción de 0,5 mg/l) generaron diferencias significativas en el porcentaje de explantes con raíz (Tabla 1).
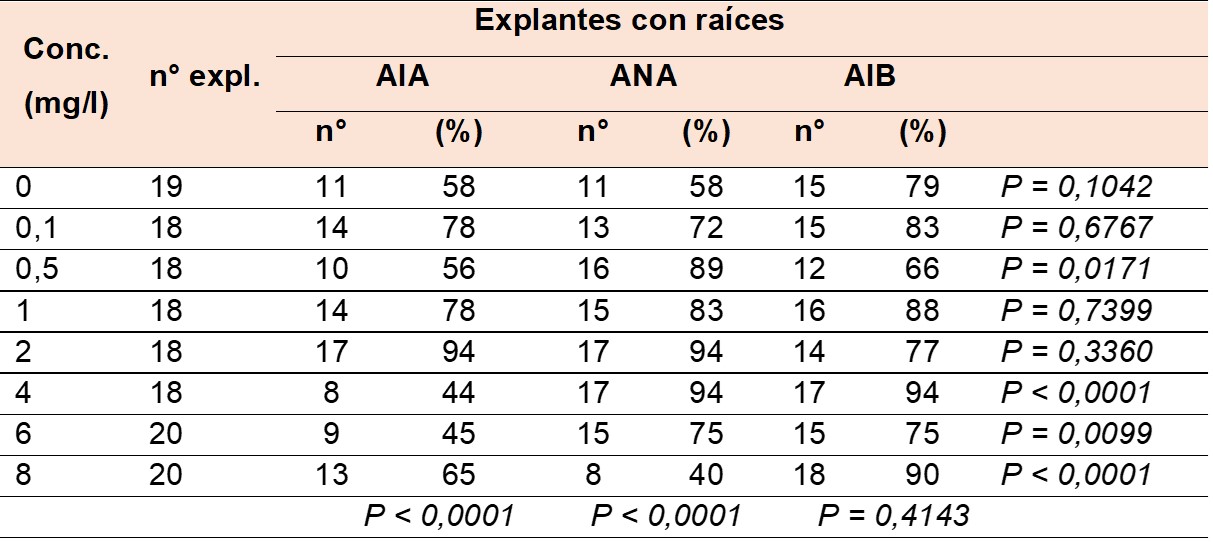
Conc. Concentraciones, n° expl. número de explantes. Los valores de P en la última columna indican pruebas de X2 para comparaciones entre auxina para una misma concentración, mientras que aquellos en la última fila indican comparaciones entre concentraciones para una misma auxina.
Conc. Concentrations, n° expl. number of explants. The P values in the last column indicate X2 tests for comparisons between auxins for the same concentration, while those in the last row indicate comparisons between concentrations for the same auxin
En relación con la variable número de raíces (NR) (Tabla 2), se observó una interacción significativa entre las auxinas y sus concentraciones según el análisis del MLGM (P < 0,0001). Las mejores respuestas fueron observadas al utilizar AIB como regulador de crecimiento, en concentraciones que oscilan de 2 a 6 mg/l. Concentraciones superiores o inferiores a este rango provocaron una disminución en el número de raíces. La formación de raíces fue promovida por ANA en un amplio rango de concentraciones, desde 0,5 hasta 6 mg/l. Respecto al AIA, los valores más altos en número de raíces formadas por explante se observaron con una concentración de 2 mg/l, seguido de 1 mg/l, notándose que concentraciones distintas a las mencionadas no resultaron en un incremento en el número de raíces por explante.
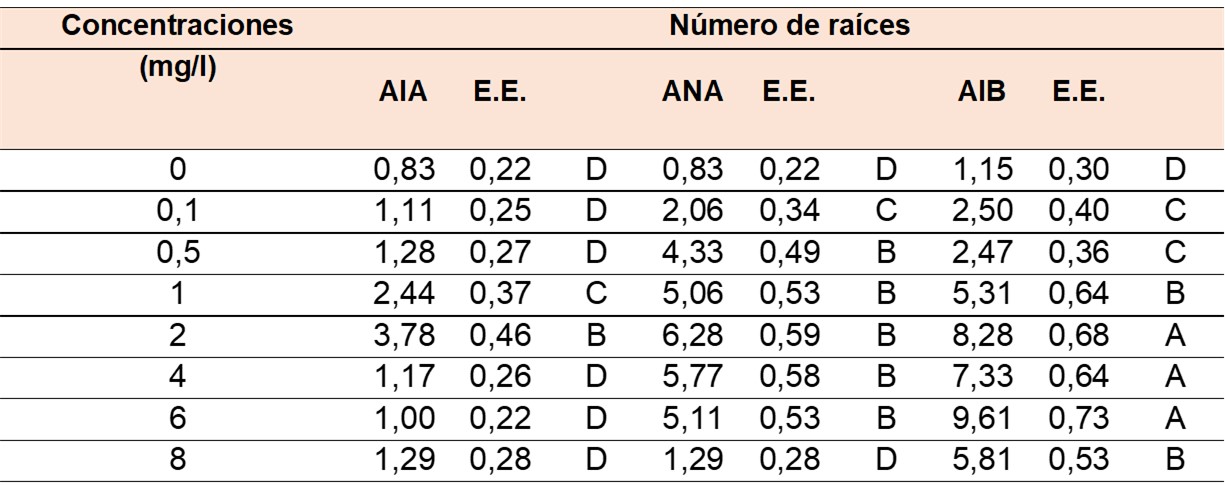
E.E. Error estándar. Medias con una letra común no son significativamente diferentes (P > 0,05).
E.S. Standard Error. Means with a common letter are not significantly different (P > 0,05).
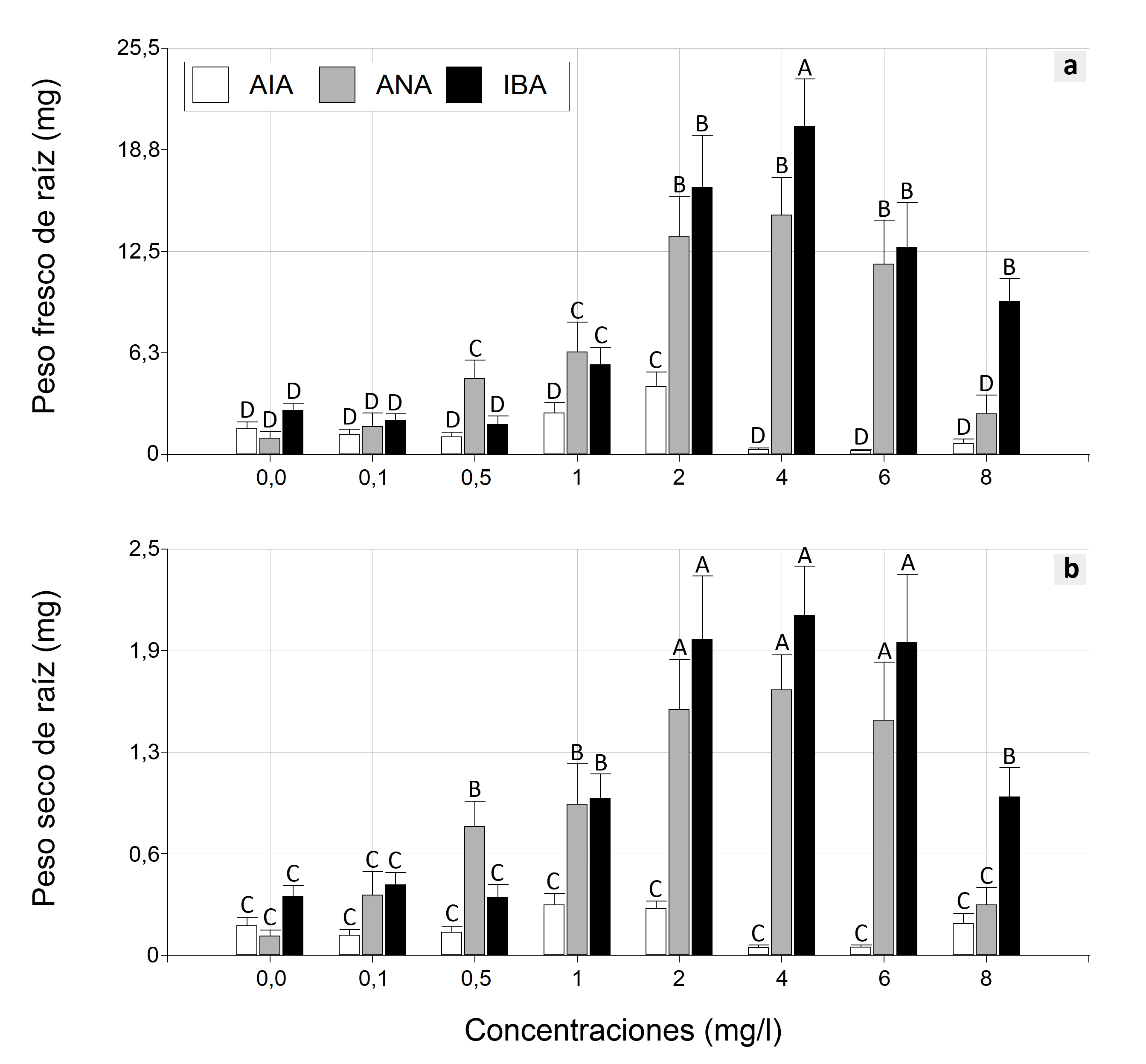
Para la variable peso fresco de raíz (PFR), el análisis del MLG mostró una interacción significativa entre las auxinas y sus concentraciones (P < 0,0001) (Figura 2.a). Se observó que la mejor respuesta en el PFR se obtuvo con una concentración de 4 mg/l de AIB. En cuanto al ANA, las respuestas más favorables se observaron en un rango de concentraciones que va de 2 a 6 mg/l. En el caso específico de AIA, se observó que solo el tratamiento con una concentración de 2 mg/l difirió significativamente del grupo de control.
También se pudo observar una interacción significativa entre las auxinas y sus concentraciones para la variable PSR (P < 0,0001). Las mejores respuestas en el PSR se obtuvieron con concentraciones de 2, 4 y 6 mg/l tanto de AIB como de ANA (Figura 2.b). Se observó una respuesta menor con las concentraciones de 1 y 8 mg/l de AIB, y con dos de los tratamientos de ANA (1 y 0,5 mg/l). Las demás concentraciones de ANA (0,1 y 0,8 mg/l) e IBA (0,1 y 0,5 mg/l), e incluso todas las concentraciones de AIA, no mostraron un efecto positivo en la variable PSR en comparación con el grupo de control.
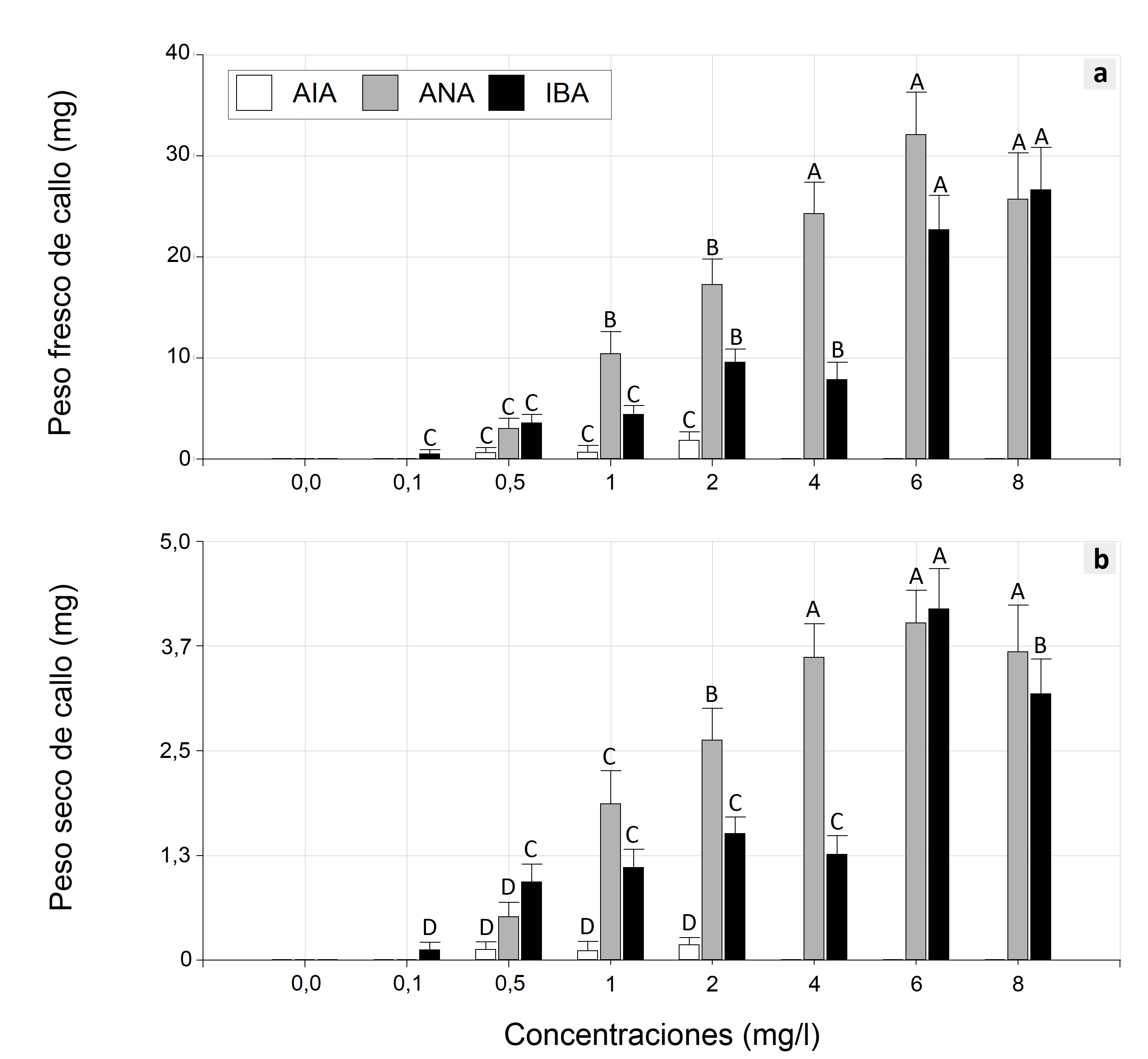
Al analizar el efecto de las auxinas en el desarrollo de callos, se observaron diferencias significativas para la interacción entre las auxinas y sus concentraciones en las variables peso fresco de callos (PFC) (P < 0,0001) y peso seco de callos (PSC) (P < 0,0001). El mayor PFC y PSC fueron producidos por el ANA y el AIB como auxinas (Figura 3.a y 3.b). Además, se pudo observar que los explantes no generaron callos en el medio de cultivo sin el agregado de reguladores de crecimiento.
Además de la formación excesiva de callos, es importante mencionar que se observaron malformaciones en el desarrollo de las raíces adventicias con concentraciones iguales o superiores a 4 mg/l de AIB y ANA. Específicamente, se observó la detención del crecimiento en las raíces, y en ocasiones se detectaron grietas o rajaduras en el tejido superficial de la raíz (Figura 4.a, 4.b y 4.c). Sin embargo, durante el ensayo con la concentración de auxina de 4 mg/l de AIB, con un tiempo de exposición de solo 7 días, seguido por el subcultivo de los explantes en el mismo medio, pero sin la auxina, se observó una disminución en la formación de callos y una reducción de las malformaciones en las raíces adventicias (Figura 4.d y 4.e). Con esta metodología de trabajo, cuando las plántulas enraizadas fueron trasladadas a un invernadero y trasplantadas, se obtuvo una tasa de sobrevivencia del 75% de las plantas (15 plántulas de 20 trasplantadas).

DISCUSIÓN
En el presente trabajo observamos que los explantes jóvenes de la muestra extraída de la población de N. alba provenientes de Campo Durán han demostrado la capacidad de regenerar raíces adventicias in vitro. Los resultados obtenidos indican que las mismas se desarrollan en presencia o ausencia de auxinas en el medio de cultivo. Observaciones similares se han realizado en segmentos uninodales de material juvenil de N. chilensis cultivados in vitro (Arce y Balboa, 1991; Caro et al., 2002). En N. alba también se ha mencionado el desarrollo de raíces en miniestacas en el tratamiento sin auxinas (de Souza et al., 2021), y de Souza y colaboradores (2014) mencionaron que la respuesta estaba asociada al estado juvenil del material a propagar. Por lo tanto, es probable que la capacidad rizogénica observada en los explantes en el medio sin auxinas se deba al estado juvenil del material vegetal utilizado para la propagación
Asimismo, se observó una respuesta positiva en la mayoría de las variables analizadas (porcentaje de explantes con raíz, NR, PFR y PSR) a medida que se aumentaban las concentraciones de auxinas en el medio, en comparación con el control. De hecho, los tres reguladores de crecimiento utilizados en el presente estudio son comúnmente los empleados para promover raíces adventicias en varias especies del mismo género (Arce y Balboa, 1991; Caro et al., 2002; Jordan et al., 1985; Tabone et al., 1986). Es importante destacar que, a pesar de que diversos estudios han señalado la actividad auxínica del 2,4-D en N. laevigata (Arya y Shekhawat, 1986; Buendía-González et al., 2007), este regulador de crecimiento no fue incluido en el presente estudio, debido a su reconocido papel como inductor de embriogénesis somática (Boeri y Sharry, 2018; Buendía-González et al., 2012) y no coincide con el objetivo de nuestro estudio.
Estos resultados demuestran que las auxinas incorporadas en el medio de cultivo estimulan la respuesta rizogénica. De las tres auxinas utilizadas, las respuestas más favorables en las variables NR, PFR y PSR se obtuvieron con ANA y AIB. En N. chilensis . N. juliflora sucede algo similar en la respuesta rizogénica cuando se utilizan ambos reguladores de crecimiento (Caro et al., 2002; Nandwani y Ramawat, 1991). Además, observamos que las concentraciones que van de 2 a 6 mg/l, tanto el ANA como el AIB mostraron respuestas similares en el PSR. Sin embargo, las concentraciones de 2, 4 y 6 mg/l de AIB resultaron en el mayor NR. Estos resultados sugieren al AIB como la auxina que induce una mejor respuesta rizogénica y concuerda con lo observado en otras especies del mismo género (Batchelor et al., 1989; Caro et al., 2002; Rivera Curi et al., 2020).
Un aspecto adicional para tener en cuenta es que tanto el AIB como el ANA demostraron ser las auxinas que indujeron el mayor desarrollo de callos (PFC y PSC). Sin embargo, las estructuras resultantes no mostraron las características típicas de callos embriogénicos descritos en Neltuma sp., que suelen ser friables, de tonalidades oscuras o verdosos, con estructuras nodulares y globulares (Boeri y Sharry, 2018; Buendía-González et al., 2012). En este estudio, los callos observados eran blanquecinos y mostraban un crecimiento celular desorganizado, probablemente debido a concentraciones supra-óptimas o a la prolongada exposición del explante a las auxinas en el medio de cultivo (De Klerk et al., 1997; Maynard y Bassuk, 1991).
Otra posible explicación de los síntomas observados, como la detención del crecimiento en las raíces o la presencia de grietas y rajaduras en el tejido superficial, podría estar relacionada con la hiperhidricidad o vitrificación (Hazarika, 2006). Aunque este fenómeno se suele observar más a menudo en brotes y tejidos aéreos de plantas cultivadas in vitro, también puede afectar a las raíces (Polivanova y Bedarev, 2022). Por ende, para comprender mejor estas malformaciones en las raíces, se deben realizar nuevos trabajos tendientes a evaluar los desórdenes morfológicos y fisiológicos asociados al tiempo de exposición a los reguladores de crecimiento en el medio de cultivo. De hecho, cuando los explantes fueron expuestos solo durante 7 días a 4 mg/l de AIB y luego transferidos a un medio sin la presencia de la auxina, se observó un desarrollo normal de las raíces y una reducción en la formación de callo.
Al trasplantar las plántulas obtenidos de explantes expuestos durante 7 días a 4 mg/l de AIB, se logró una tasa de supervivencia del 75%. Estos resultados son similares con trabajos previos en especies del mismo género, especialmente cuando se emplean explantes jóvenes o plántulas provenientes de viveros (Arce y Balboa, 1991; Goyal y Arya, 1984; Jordan et al., 1985; Tabone et al., 1986).
CONCLUSIONES
Este estudio demuestra que los explantes jóvenes de N. alba son capaces de desarrollar raíces adventicias cuando son cultivados in vitro. Se observa una mayor respuesta rizogénica a medida que aumentan las concentraciones de auxinas. Tanto el ANA como el AIB muestran las mejores respuestas en las variables analizadas, especialmente en concentraciones que van de 2 a 6 mg/l. El subcultivo de los explantes en un medio sin auxinas, después de 7 días de exposición a 4 mg/L de IBA, ha demostrado reducir la formación de callos y minimizar las malformaciones en las raíces adventicias. Esta metodología de trabajo permite alcanzar una tasa de sobrevivencia del 75% de las plantas ex vitro.
BIBLIOGRAFÍA
Arce, P., y Balboa, O. (1991). Seasonality in rooting of Prosopischilensis cuttings and in-vitro micropropagation. Forest Ecology and Management, 40(3-4), 163-173. https://doi.org/10.1016/0378-1127(91)90036-U
Arya, H. C., y Shekhawat, N. S. (1986). Clonal multiplication of tree species in the Thar desert through tissue culture. Forest ecology and management, 16(1-4), 201-208. https://doi.org/10.1016/0378-1127(86)90020-4
Batchelor, C. A., Yao, D., Koehler, M. J., y Harris, P. J. C. (1989). In vitro propagation of Prosopis species .P. chilensis, P. cineraria and P. juliflora). In Annales des Sciences Forestieres (Vol. 46, No. Supplement, pp. 110s-112s). EDP Sciences.
Boeri, P., y Sharry, S. (2018). Somatic embryogenesis of alpataco (Prosopis alpataco L.). Step Wise Protocols for Somatic Embryogenesis of Important Woody Plants: Volume II, 189-198. https://doi.org/10.1007/978-3-319-79087-9_16
Brondani, G. E., de Wit Ondas, H. W., Baccarin, F. J. B., Gonçalves, A. N., y de Almeida, M. (2012). Micropropagation of Eucalyptus benthamii to form a clonal micro-garden. In Vitro Cellular & Developmental Biology-Plant, 48, 478-487. DOI 10.1007/s11627-012-9449-9
Buendía-González, L., Orozco-Villafuerte, J., Cruz-Sosa, F., Chávez-Ávila, V. M., y Vernon-Carter, E. J. (2007). Clonal propagation of mesquite tree (Prosopislaevigata Humb. & Bonpl. ex Willd. MC Johnston). I. Via cotyledonary nodes. In Vitro Cellular & Developmental Biology-Plant,43, 260-266. DOI 10.1007/s11627-007-9027-8
Buendía-González, L., Estrada-Zúñiga, M. E., Orozco-Villafuerte, J., Cruz-Sosa, F., y Vernon-Carter, E. J. (2012). Somatic embryogenesis of the heavy metal accumulator Prosopis laevigata. PlantCell, Tissue and Organ Culture, 108, 287-296. DOI 10.1007/s11240-011-0042-4
Caro, L. A., Polci, P. A., Lindström, L. I., Echenique, C. V., y Hernandez, L. F. (2002). Micropropagation of Prosopis chilensis (Mol.) Stuntz from young and mature plants. BIOCELL, 26(1), 25-33.
Castillo de Meier, G., y Bovo, O. A. (2000). Plant regeneration from single-nodal-stem explants of legume tree Prosopis alba Griseb. Biocell: Official Journal of the Sociedades Latinoamericanas de Microscopia Electronica, 24(2), 89-95.
De Klerk, G. J., Brugge, J. T., y Marinova, S. (1997). Effectiveness of indoleacetic acid, indolebutyric acid and naphthaleneacetic acid during adventitious root formation in vitro in Malus ‘Jork 9’. Plant cell, tissue and organ culture, 49(1), 39-44.
de Souza, J. C. A. V., Bender, A. G., Tivano, J. C., Barroso, D. G., Mroginski, L. A., Vegetti, A. C., y Felker, P. (2014). Rooting of Prosopis alba mini-cuttings. New Forests, 45, 745-752. DOI 10.1007/s11056-014-9429-5
de Souza, J. C. A. V., Bender, A. G., Reutemann, A. G., Perreta, M. G., Córdoba, M. S., Tivano, J. C., Barroso, D. G., Gariglio, N. F., Mroginski, L. A y Vegetti, A. V. (2021). Influencia del grado de lignificación de los propágulos de jardín y minijardín clonal en el enraizamiento de estacas y miniestacas de Prosopis alba Griseb. FAVE Sección Ciencias Agrarias, 20(1), 287-304. DOI 10.14409/fa.v20i1.10270
Di Rienzo J. A., Guzmán A.W., y Casanoves F. (2002). A multiple comparisons method based on the distribution of the root node distance of a binary tree. Journal of Agricultural, Biological, and Environmental Statistics, 7,1-14.
Di Rienzo, J. A., Casanoves F., Balzarini M. G., Gonzalez L., Tablada M., y Robledo C. W. (2012). InfoStat. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. http://www.infostat.com.ar.
Goyal, Y., y Arya, H. C. (1984). Tissue culture of desert trees: I. Clonal multiplication of Prosopis cineraria by bud culture. Journalof Plant Physiology, 115(3), 183-189. https://doi.org/10.1016/S0176-1617(84)80118-2
Green, B., Tabone, T., y Felker, P. (1990). A comparison of amide and ureide nitrogen sources in tissue culture of tree legume Prosopisalba clone B 2 V 50. Plant cell, tissue and organ culture, 21, 83-86.
Hartmann, H.T.; D.E. Kester; y R.L. Geneve. (2011). Plant propagation principles and practices Prentice Hall.
Hazarika, B. N. (2006). Morpho-physiological disorders in in vitro culture of plants. Scientia Horticulturae, 108(2), 105-120. https://doi.org/10.1016/j.scienta.2006.01.038
Hughes, C. E., Ringelberg, J. J., Lewis, G. P., y Catalano, S. A. (2022). Disintegration of the genus Prosopis L. (Leguminosae, Caesalpinioideae, mimosoid clade). PhytoKeys, 205, 147. doi: 10.3897/phytokeys.205.75379
Jordan, M. (1987). In vitro culture of Prosopis species. In Cell and Tissue Culture in Forestry: Case Histories: Gymnosperms, Angiosperms and Palms (pp. 370-384). Dordrecht: Springer Netherlands.
Jordan, M., Pedraza, J., y Goreux, A. (1985). In vitro propagation studies of three Prosopis species (P. alba, P. chilensis and P. tamarugo) through shoot-tip culture. Gartenbauwissenschaft, 50(6), 265-267.
Maynard, B. K., y Bassuk, N. L. (1991). Stock plant etiolation and stem banding effect on the auxin dose-response of rooting in stem cuttings of Carpinusbetulus L.‘Fastigiata’. Plant growth regulation, 10, 305-311. https://doi.org/10.1007/BF00024590
Minchala-Patiño, J., Poma-Angamarca, R., Muñóz-Chamba, L., Yaguana-Arévalo, M., González-Zaruma, D., Eras-Guamán, V. H., C. Rojas-Idrogo, G. E. y Delgado-Paredes, G. E. (2014). Propagación in vitro de Prosopis limensis Benth. in Hook. (Fabaceae-Mimosoideae). Quebracho(Santiago del Estero), 22(2), 88-99.
Morales-Domínguez, J. F., de León, D. S. D., Garcidueñas-Piña, C., y Pérez-Molphe-Balch, E. (2019). Germination, in vitro propagation and soil acclimatization of Acacia farnesiana and Prosopislaevigata. South African Journal of Botany, 124, 345-349. https://doi.org/10.1016/j.sajb.2019.05.034
Murashige, T., y Skoog, F. (1962). A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant, 15, 473-497.
Nandwani, D., y Ramawat, K. G. (1991). Callus culture and plantlets formation from nodal explants of Prosopis juliflora (Swartz) DC. Indian J Exp Biol, 29(6), 523-527.
Polivanova, O. B., y Bedarev, V. A. (2022). Hyperhydricity in plant tissue culture. Plants, 11(23), 3313. https://doi.org/10.3390/plants11233313
Rivera Curi, J. C., Cabrera Pintado, R. M., y Bulnes Soriano, F. (2020). Micropropagación de Prosopis pallida (Humb &Bonpl. Ex Willd.) Kunth a partir de yemas apicales. Revista Colombiana de Biotecnología, 22(1), 18-26.
Shekhawat, N. S., Rathore, T. S., Singh, R. P., Deora, N. S., y Rao, S. R. (1993). Factors affecting in vitro clonal propagation of Prosopiscineraria. Plant Growth Regulation, 12, 273-280. https://doi.org/10.1007/BF00027208
Tabone, T. J., Felker, P., Bingham, R. L., Reyes, I., y Loughrey, S. (1986). Techniques in the shoot multiplication of the leguminous tree Prosopis alba clone B2V50. Forest ecology and management, 16(1-4), 191-200. https://doi.org/10.1016/0378-1127(86)90019-8
Vega, C., Aguilar, D., Bessega, C., Teich, I., Acosta, M. C., Cosacov, A., Ewens, M., Vilardi, J., A. N. Sérsic y Verga, A. (2021). Genetic Basis for their Domestication and Conservation. Genetic Variation Patterns of “Algarrobos” from the “Great American Chaco” (Prosopisalba, P. nigra, P. hassleri, P. fiebrigii, P. ruscifolia, P. chilensis, and P. flexuosa). En: M. J. Pastorino & P. Marchelli (Eds.). Low Intensity Breeding of Native Forest Trees in Argentina. Springer. pp. 245-269. https://doi.org/10.1007/978-3-030-56462-9_9
Venier, M. P., Cosacov Martinez, A., López Lauenstein, D., Vega, C., y Verga, A. (2013). Impacto del cambio climático sobre la distribución de Prosopishassleri y P. alba en la región Chaqueña. Producción Forestal, 7, 35-38.
Venier, P., Ferreras, A. E., Verga, A., y Funes, G. (2015). Germination traits of Prosopis alba from different provenances. Seed Science and Technology, 43(3), 548-553. https://doi.org/10.15258/sst.2015.43.3.14
Verdes, P. (2007). Notas: micropropagación de Prosopis caldenia burk.: estado actual y perspectivas. Revista Científica Agropecuaria, 11(1), 45-51.
Verga, A., y López Lauenstein, D. (2021). Genetic Basis for their Domestication and Conservation. Subtropical dry forests: the main forest ecoregion of Argentina. En: M. J. Pastorino & P. Marchelli (Eds.), Low Intensity Breeding of Native Forest Trees in Argentina. Springer. pp. 217-243. https://doi.org/10.1007/978-3-030-56462-9

