Artículos
Cambios en propiedades del suelo y su influencia en la macrofauna en secuencias de cultivos que incluyen cultivos de cobertura
Changes in soil properties and their influence on macrofauna in crop sequences including cover crops
 Sergio Montico smontico@unr.edu.ar
Guillermo Alberto Montero gmontero@unr.edu.ar
Sergio Montico smontico@unr.edu.ar
Guillermo Alberto Montero gmontero@unr.edu.ar
Revista FAVE Sección Ciencias Agrarias
Universidad Nacional del Litoral, Argentina
ISSN: 2346-9129
ISSN-e: 2346-9129
Periodicidad: Semestral
núm. 23, e0029, 2024
Recepción: 28 Octubre 2023
Aprobación: 24 Mayo 2024
Autor de correspondencia: scaglionejosefina@gmail.com

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional.
Resumen: Los objetivos de este trabajo fueron evaluar el efecto de los cultivos de cobertura (CC) y las secuencias de cultivos sobre algunas propiedades del suelo y la abundancia de la macrofauna edáfica; y la influencia de estas propiedades sobre la presencia de las principales familias de macroinvertebrados. Se trabajó en un experimento instalado en Zavalla (Santa Fe) cuyos tratamientos incluyeron CC de vicia y triticale como antecesores de maíz y soja, respectivamente, además de dos tratamientos de barbecho con los mismos cultivos de cosecha sucesores. Los muestreos se realizaron después del ciclo de los CC (Mcc-b) y de los cultivos de cosecha (Mf). Entre las propiedades edáficas, el carbono orgánico particulado registró un incremento en las secuencias que incluyeron CC, en tanto que la resistencia mecánica a la penetración y el porcentaje de agregados estables al agua presentaron variaciones a corto plazo que estuvieron vinculadas con la actividad de las raíces de los CC, particularmente luego de la supresión de la vicia. Por su parte, los cambios en la abundancia de macrofauna se asociaron con las condiciones edáficas generadas por los distintos tratamientos. En el Mcc-b existió una diferenciación entre ambos CC, y de éstos con respecto al barbecho, en tanto que en el Mf las muestras que incluyeron CC mostraron condiciones edáficas más favorables respecto a las del barbecho. Finalmente, los CC indujeron la presencia de organismos de la macrofauna pertenecientes a distintos grupos tróficos debido a la cobertura generada y al aporte de recursos alimenticios.
Palabras clave: Hábitat del suelo, macroinvertebrados, condición físico-mecánica, indicadores de salud edáfica.
Abstract: The objectives of this study were to evaluate the effects of cover crops (CC) and crop sequences on some soil properties and macrofauna abundance; and the influence of these properties on the presence of the main macroinvertebrates’ families. We worked on an experiment installed in Zavalla (Santa Fe) which treatments included CC of vetch and triticale before maize and soybean, respectively, and two treatments of fallow with the same cash crops. Samplings were done after CC (Mcc-b) and cash crops (Mf) cycles. Among edaphic properties, particulate organic carbon registered an increase in sequences with CC, while mechanical penetration resistance and water stable aggregates percentage showed short term variations that were linked with CC´s root development, particularly after vetch suppression. Changes in the abundance of macrofauna were associated with edaphic conditions generated by the treatments. In Mcc-b there was a differentiation between samples of both CC, and between them and fallow ones, while in Mf samples which included CC showed more favorable edaphic conditions in relation with fallow ones. Finally, CC leaded the presence of macrofaunal organisms from different trophic groups due to the coverage generated and the input of food resources.
Keywords: Soil habitat, macroinvertebrates, physical-mechanical condition, soil health indicators.
Introducción
La intensificación sustentable de las secuencias rotacionales juega un rol crucial en el desarrollo de agroecosistemas que alcancen los niveles de producción deseados en consonancia con la conservación de los recursos. En particular, la conservación de los suelos y el mantenimiento de su condición de salud resulta de relevancia ya que los mismos proveen una gran variedad de hábitats que albergan distintos tipos de organismos y de los cuales depende la provisión de servicios ecosistémicos (FAO, 2020). En este sentido, las plantas influyen directamente sobre la configuración del hábitat edáfico y, por ende, en los organismos que residen en él (Geisen et al., 2019). Esto se vincula con las modificaciones que generan sobre la condición física y química del suelo debido al aporte de diferente cantidad y calidad de residuos y al crecimiento de las raíces (Berg y Smalla, 2009). Además, la vegetación interviene directamente sobre la presencia de organismos herbívoros, como así también de patógenos y simbiontes (Sylvain y Wall, 2011). Por lo tanto, en ecosistemas agrícolas el manejo de la vegetación (cultivable o no) y los residuos que ésta aporta resultan, en conjunto con otros factores y prácticas de manejo, los principales reguladores del hábitat edáfico y de los organismos que se desarrollan en él (Laossi et al., 2008). Entre ellos, la macrofauna posee relevancia en el funcionamiento del suelo debido a su intervención en la modificación de la estructura, la descomposición de la materia orgánica y el ciclado de nutrientes, entre otros aspectos (Wagg et al., 2014).
Los cultivos de cobertura (CC) han sido identificados como alternativas de intensificación de las secuencias rotacionales ya que contribuyen a incrementar la resiliencia del sistema suelo frente a distintos disturbios (White et al., 2016). Esto se vincula con sus múltiples efectos, entre los que se destacan la reducción de las pérdidas de nutrientes, el incremento del carbono orgánico del suelo, los cambios en la abundancia y diversidad de la fauna edáfica y la reducción en la dependencia del sistema hacia insumos externos (Crotty y Stoate, 2019). Asimismo, en lo que respecta a la fauna del suelo, se ha demostrado que el impacto positivo de los CC sobre sus comunidades se asocia con cambios en el hábitat edáfico y en las redes tróficas que allí se desarrollan (Kelly et al., 2021).
Los objetivos de este trabajo fueron: 1) evaluar el efecto de los CC y la configuración de las secuencias de cultivos sobre algunas propiedades del suelo y la abundancia de organismos de la macrofauna; 2) interpretar la influencia de las propiedades edáficas seleccionadas sobre la presencia de las principales familias de la macrofauna.
Materiales y Métodos
Experimento
El estudio se llevó a cabo durante la campaña 2020/2021, en un experimento instalado en un lote perteneciente al Campo Experimental J. V. Villarino, ubicado en la localidad de Zavalla (Santa Fe, Argentina) (33º01´S; 60º53´O). El suelo correspondió a un complejo constituido por las series Roldán (70%) (Argiudol vértico), Monte Flores (20%) (Argialbol típico) y Gelly (10%) (Natralbol típico) (INTA, 2023).
El experimento se desarrolló en un lote con historia productiva agrícola de al menos diez años. El diseño fue en bloques completos aleatorizados con tres repeticiones y las unidades experimentales fueron parcelas de 20 m x 20 m en las que se evaluaron secuencias de cultivos con y sin inclusión de CC durante el período otoño-invernal y diferentes cultivos de cosecha sucesores. El experimento se inició a principios de mayo de 2020 con la siembra de los CC y se prolongó hasta la cosecha de los cultivos estivales en mayo de 2021. Durante ese período, el monto acumulado de precipitaciones fue de 811 mm y la temperatura media de 16,9 ºC, según los registros de la estación agrometeorológica del Campo Experimental J. V. Villarino.
Los tratamientos analizados fueron: 1) barbecho – maíz (Zea mays) (Bar-M); 2) CC vicia (Vicia villosa R.) (CCv) – maíz (CCv-M); 3) barbecho – soja (Glycine max (L.) Merr.. (Bar-S); 4) CC triticale (x Triticosecale W.) (CCt) – soja (CCt-S). Todas las secuencias presentaron antecesor trigo (Triticum aestivum) – soja.
El CCv se sembró el 08/05/2020 (28 kg ha-1) y el CCt el 11/05/2020 (90 kg ha-1) empleando una sembradora de siembra directa con distancia entre surcos de 19 cm. En ese momento se aplicó un fertilizante con 7% de nitrógeno, 40% de fósforo, 9% de azufre y 10% de magnesio en dosis de 75 kg ha-1 para ambos CC. El ciclo de los mismos se interrumpió de manera química el 05/11/2020 empleando glifosato con una dosis de 2 L ha-1. La siembra de los cultivos estivales sucesores se efectuó el 04/11/2020 en el caso del maíz (DM 4615 STS) y el 06/11/2020 en soja (22.6 PWU), para lo cual se utilizó una sembradora de siembra directa con distancia entre surcos de 52 cm. La densidad de siembra del maíz fue de 4 semillas por metro lineal, mientras que en soja fue de 22 semillas por metro lineal. En todos los tratamientos se realizó una fertilización a la siembra con fosfato monoamónico (11-52-0) en una dosis de 80 kg ha-1. El control de malezas a lo largo del experimento se efectuó de manera química, realizando las pulverizaciones según estado y densidad de las mismas, en tanto que en ninguna oportunidad a lo largo del período evaluado fue necesario aplicar insecticidas y fungicidas. La cosecha de soja se realizó el 21/04/2021, mientras que la de maíz se llevó a cabo el 29/04/2021.
Muestreos
Los muestreos se realizaron en dos oportunidades: al momento de la supresión del crecimiento de los CC y antes de la siembra de los cultivos de cosecha (02/11/2020) (Mcc-b), y luego de la cosecha de los cultivos de maíz y soja (11/05/2021) (Mf). Dentro de cada parcela se definió un cuadrado de 6 m de lado en cuyos vértices se extrajeron las muestras para análisis de macrofauna y en el centro las correspondientes a variables edáficas. El espesor de suelo estudiado fue 0-20 cm.
Variables evaluadas
Las propiedades edáficas analizadas fueron: carbono orgánico total (COT, %), carbono orgánico particulado (COP, %), agregados estables al agua (AEA, %) y resistencia mecánica a la penetración (RMP, KPa). Para las dos primeras, en cada parcela se extrajo una muestra compuesta constituida por 12 a 15 submuestras. El COT se obtuvo mediante el método de calcinación (Chatterjee et al., 2009) y el COP se determinó a través de la separación física de las muestras (Cambardella y Elliot, 1992) y el método Walkley Black (Walkley y Black, 1934) para la estimación del contenido de carbono. En este caso, el COP correspondió a la fracción retenida en el tamiz de 53 µm de tamaño de malla. El porcentaje de AEA se determinó según la metodología descripta por Hénin et al. (1958) y considerando la fracción de agregados comprendida entre 0,25 y 2 mm. La RMP se midió empleando un penetrómetro digital Fieldscout SC 900 y se efectuó la corrección de los valores al 18% de humedad edáfica según la ecuación propuesta por Krüger et al. (2018).
Para el estudio de la macrofauna, en cada parcela se extrajeron cuatro monolitos de suelo de 20 cm de lado por 20 cm de profundidad, en los cuales se separaron manualmente los organismos presentes (Adaptado de Anderson e Ingram, 1993). Los ejemplares colectados en cada muestra fueron colocados en solución conservante de alcohol 70% V/V o formaldehído 5% V/V en el caso de las lombrices, y determinados hasta el nivel de familia empleando las correspondientes claves taxonómicas (Borror et al., 1992; Morrone y Coscarón, 1998; Roig-Juñent et al., 2014a, 2014b; Stehr, 1991). En cada muestra se cuantificó la abundancia por familia mediante conteo manual.
Análisis estadísticos
Las diferencias entre tratamientos y momentos de muestreo en las propiedades edáficas se evaluaron mediante Modelos Lineales Generales y Mixtos (Di Rienzo et al., 2011). En los mismos se consideró el tratamiento, momento de muestreo, bloque y la interacción entre los dos primeros como efectos fijos, mientras que la parcela fue el efecto aleatorio. El análisis de la abundancia de organismos de la macrofauna se realizó a través de Modelos Lineales Generalizados y Mixtos (Di Rienzo et al., 2017), considerando los mismos efectos fijos y aleatorio anteriores y distintas familias de distribución (Poisson y Binomial Negativa). Del conjunto de modelos planteados en cada variable se seleccionó el que presentó el valor más bajo de los criterios de Akaike (AIC) y Bayesiano de Información (BIC) y, en el caso de la abundancia de macrofauna, también se consideró que el cociente entre la deviance del modelo y los grados de libertad residuales fuese cercano a uno. Cuando existieron diferencias significativas entre factores, las mismas se evaluaron mediante la prueba LSD Fisher (α=0,05). Los análisis se realizaron a través de la interfaz de R incluida en el software InfoStat (Di Rienzo et al., 2020).
Se ejecutaron Análisis de Correspondencia Canónica (ACC) (Ter Braak, 1986) para cada momento de muestreo considerando los datos de las propiedades edáficas y la abundancia de las familias de macrofauna con mayor representatividad en cada caso (abundancia superior a 10 organismos). Los análisis se efectuaron con el software PCORD v6.0 (Mc Cune y Mefford, 2011).
Resultados y Discusión
Propiedades edáficas
En cuanto a las propiedades edáficas (Tabla 1), sólo existió interacción significativa entre tratamientos y momentos de muestreo en RMP (F = 5,80; p = 0,007), en tanto que el COT y el porcentaje de AEA mostraron diferencias significativas entre momentos de muestreo (COT: F = 17,44; p = 0,001; Porcentaje de AEA: F = 6,87; p = 0,02). Por su parte, el contenido de COP sólo registró diferencias significativas entre tratamientos (F = 15,14; p = 0,0001).
Con respecto a la RMP, se detectó un valor significativamente superior asociado al CCv en el Mcc-b en relación a los restantes tratamientos. Asimismo, dicho tratamiento fue el único que registró variaciones significativas entre ambos momentos de muestreo, observándose una disminución en el Mf. El desarrollo radical más profuso detectado en el CCv en comparación con el CCt en el espesor de suelo considerado podría haber generado una densificación del suelo circundante a las raíces debido al consumo de agua y a la generación de nuevos canales entre los agregados (Bodner et al., 2014), lo cual conduciría a un incremento de la RMP. Dicha tendencia reflejaría un efecto de corto plazo de los CC sobre la propiedad en cuestión ya que, en períodos de tiempo mayores y bajo diferentes esquemas rotacionales, se ha demostrado que los CC contribuyen a reducir la RMP y, con ello, a mitigar los efectos de la compactación (Blanco-Canqui y Ruis, 2020). Por otro lado, la magnitud del impacto de los CC sobre las propiedades edáficas a lo largo del tiempo, por ejemplo, en la estructura, respondería a una serie de factores, entre los que se destacan las especies de CC consideradas, la duración del período de crecimiento y las condiciones ambientales que, en conjunto, influirían sobre la cantidad de biomasa producida (Blanco-Canqui y Jasa, 2019). Al respecto, se registró un porcentaje de AEA significativamente superior en el Mcc-b (26,67 ± 2,35 %) en comparación con el Mf (19,97 ± 1,02 %). Además, si bien no existieron diferencias significativas, en el Mcc-b el porcentaje de AEA fue mayor en los tratamientos que incluyeron CC en relación a los de barbecho, mientras que en el Mf dicha tendencia sólo se observó en la secuencia CCv-M. En este sentido, podrían notarse ciertos efectos asociados a los CC que se vinculan con su desarrollo radical y la actividad biológica (Castiglioni y Behrends Kraemer, 2019). En este punto, Hudek et al. (2022) indicaron una correlación positiva entre la estabilidad de los agregados y la densidad de raíces de distintas especies de CC que se vinculó ampliamente con la promoción de la actividad microbiana derivada de la rizodeposición de exudados, células y residuos que representan fuentes de energía para la microbiota rizosférica, la cual contribuye activamente a la agregación del suelo (Miransari, 2014).
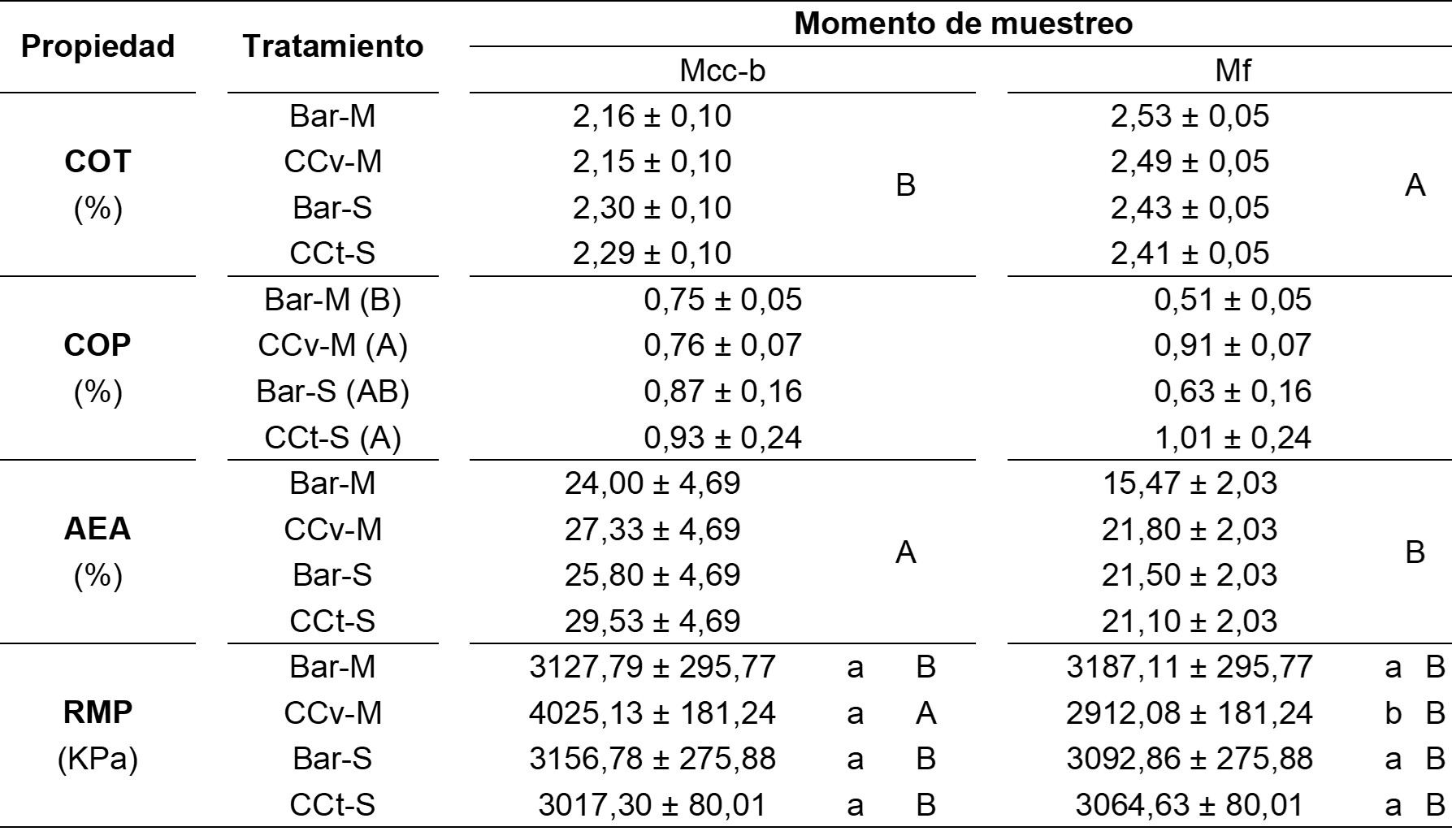
En referencia al COT, en el Mf se evidenció un contenido significativamente superior (2,47 ± 0,03 %) que en el Mcc-b (2,23 ± 0,05 %). En este caso, dado el corto período de tiempo transcurrido entre ambos muestreos, posiblemente tales diferencias no definan una tendencia consistente en cuanto a la evolución de la propiedad, debido a la multiplicidad de compuestos y fracciones que forman parte de la materia orgánica y cuya dinámica temporal es particular (Zhang et al., 2022). Por lo tanto, sería necesario contar con información derivada de la evaluación en un período de tiempo mayor y que refleje efectos acumulativos asociados a la inclusión de CC en las secuencias de cultivos (Chahal et al., 2020; Duval et al., 2016). Asimismo, el incremento del COT es altamente dependiente de la calidad y cantidad de residuos aportados por cada especie de CC (Chahal et al., 2020; Jian et al., 2020). Por el contrario, en lo que respecta al COP, existió una variación significativa entre los tratamientos Bar-M (0,63 ± 0,02 %) y CCv-M (0,83 ± 0,03 %), mientras que no se registraron cambios significativos entre ambos tratamientos de barbecho, como así tampoco entre Bar-S (0,75 ± 0,06 %) y CCt-S (0,97 ± 0,1 %). Además, si bien las diferencias no fueron significativas, en el Mf se observó que los tratamientos con CC presentaron mayor contenido promedio de COP que los de barbecho, lo cual es coincidente con lo descripto por Romaniuk et al. (2018) en situaciones con CC y barbecho en el período de un año.
Abundancia de macrofauna y vinculación con propiedades edáficas
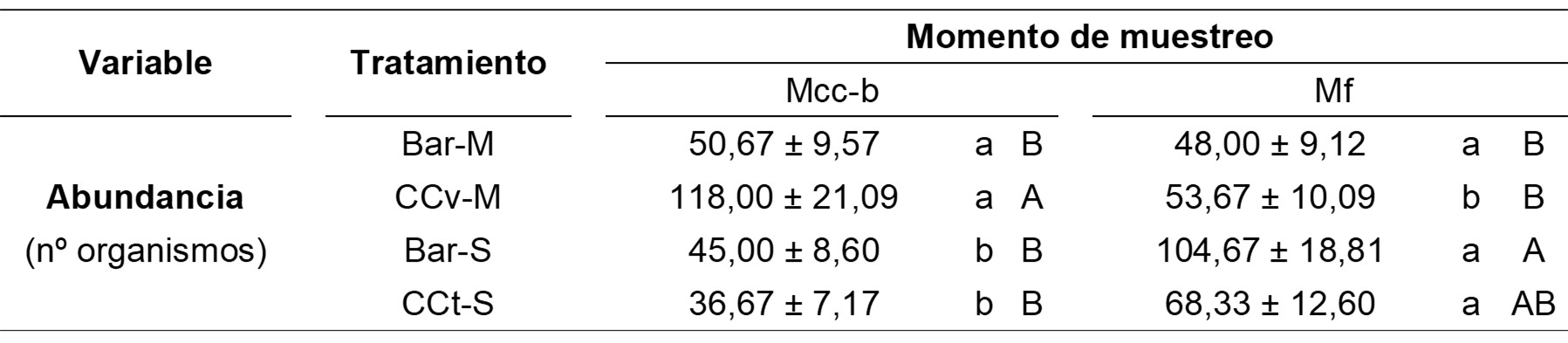
La abundancia de macrofauna fue afectada significativamente por la interacción entre tratamientos y momentos de muestreo (X2 = 7,92; . = 0,002) (Tabla 2). En el Mcc-b se observó un valor significativamente superior asociado al CCv, mientras que en el Mf la mayor abundancia correspondió a la secuencia Bar-S. Asimismo, CCt-S mostró una tendencia de incremento en el Mf en relación al Mcc-b. En este sentido, el efecto de los CC sobre la abundancia de macrofauna podría ser diferente según los grupos que la componen (Tablas 3 y 4), teniendo en cuenta que los mismos exhiben variados hábitos alimenticios y estrategias de comportamiento (Lavelle et al., 2022). Al respecto, Menta et al. (2020) propusieron que la abundancia de invertebrados del suelo usualmente resulta más afectada por el cultivo de cosecha o la configuración de las secuencias que por el manejo de los residuos o la inclusión de CC. Esto estaría en relación con que los efectos de los CC podrían resultar variables en función del período de tiempo considerado y el grado de adaptación de los organismos a las condiciones particulares del suelo, lo cual define su sensibilidad frente a los cambios en el ambiente edáfico (Fiorini et al., 2022).
En las Tablas 3 y 4 se muestra la abundancia de organismos de cada familia según momentos de muestreo y tratamientos. Se registró una mayor cantidad de familias en el Mcc-b en comparación con el Mf, pero existió una menor abundancia total en el primer muestreo que en el segundo.
En las Figuras 1 y 2 se presentan los ACC relativos a cada momento de muestreo. En el Mcc-b, el porcentaje de varianza explicado por los dos primeros ejes canónicos fue de 45 %, mientras que en el Mf dicho porcentaje fue de 36 %.
La calidad del hábitat edáfico puede ser afectada por múltiples factores, entre los que se incluyen la cantidad y calidad de recursos alimenticios, de sitios de anidamiento o refugio y la abundancia de competidores o predadores (Halliday et al., 2019). Además, se ha planteado que la diversidad de hábitats presentes en el suelo queda definida por la vegetación y las condiciones edáficas, las cuales son modificadas directamente por la primera (Stasiov et al., 2021).
En relación con ello, en el Mcc-b las muestras pertenecientes al CCv se asociaron con valores elevados de RMP y favorecieron la presencia de hormigas (Formicidae) y de distintas familias de coleópteros (Curculionidae, Staphylinidae y Tenebrionidae) (Figura 1), las cuales poseen hábitos alimenticios herbívoros, predadores y detritívoros, respectivamente.
En relación a las hormigas, se han definido variaciones en sus preferencias de hábitat debido a que presentan alta diversidad, abundancia y hábitos generalistas de explotación de recursos que les permiten ocupar una gran variedad de nichos y disponer de un amplio rango de recursos alimenticios que les posibilitan competir y sobrevivir con mayor éxito en relación con otros organismos del suelo (Chanatásig-Vaca et al., 2011). Por otro lado, las muestras correspondientes al CCt se asociaron con condiciones edáficas más favorables y que estuvieron representadas por contenidos elevados de COT y, en menor medida, con COP y porcentaje de AEA. En este caso, también se vio promovida la presencia de organismos detritívoros (Pseudonnanolenidae, Onychiuridae), herbívoros (Chrysomellidae) y predadores (Henicopidae, Carabidae). La ocurrencia de detritívoros asociada a los CC podría responder, no sólo a la disponibilidad de recursos alimenticios (residuos), sino también a que estos organismos, al habitar entre los residuos superficiales o en los primeros centímetros del suelo, son particularmente sensibles a los cambios bruscos de temperatura y humedad, los cuales serían regulados por la cobertura generada (Cabrera, 2012). Por otro lado, se ha reportado que la presencia de herbívoros responde a una serie de factores, entre los que se destacan la cantidad y calidad de los materiales vegetales disponibles (Masters, 2004) y la abundancia de organismos predadores debido a la interacción trófica existente entre ambos grupos (Murrel, 2017). En referencia a estos últimos, se ha indicado que los predadores epigeos se verían beneficiados por los sitios de refugio generados en los residuos superficiales y por la disponibilidad de presas allí presentes (Bedano et al., 2016; Jiang et al., 2018) y que, en términos generales, los CC promoverían su presencia ya que contribuyen a incrementar la complejidad del hábitat edáfico en ecosistemas agrícolas (Rosario-Lebron et al., 2018). Asimismo, la mejora en la condición físico-mecánica del suelo asociada a los CC actuaría favorablemente sobre el movimiento de los predadores hipogeos en la matriz del suelo (Erktan et al., 2020).
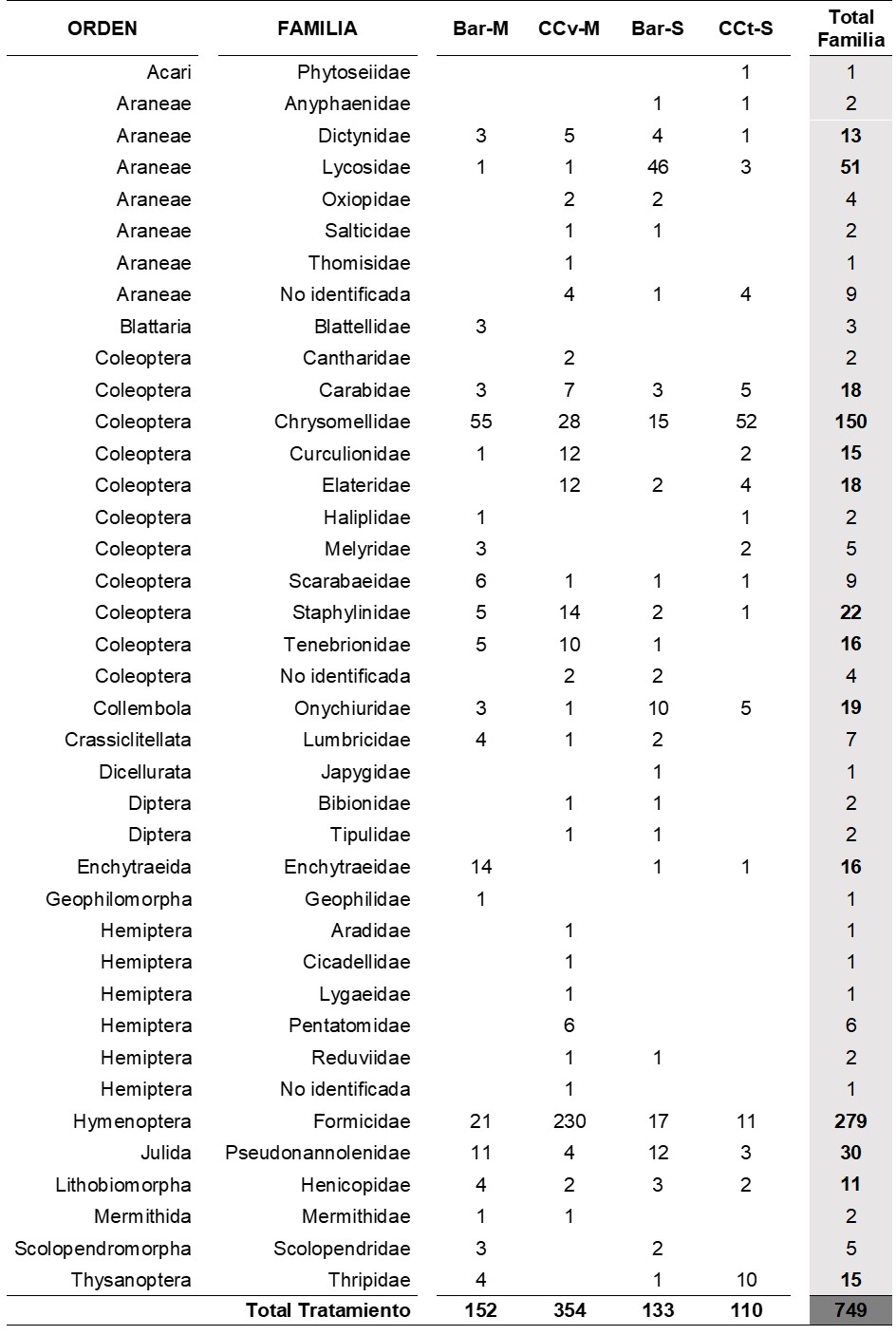
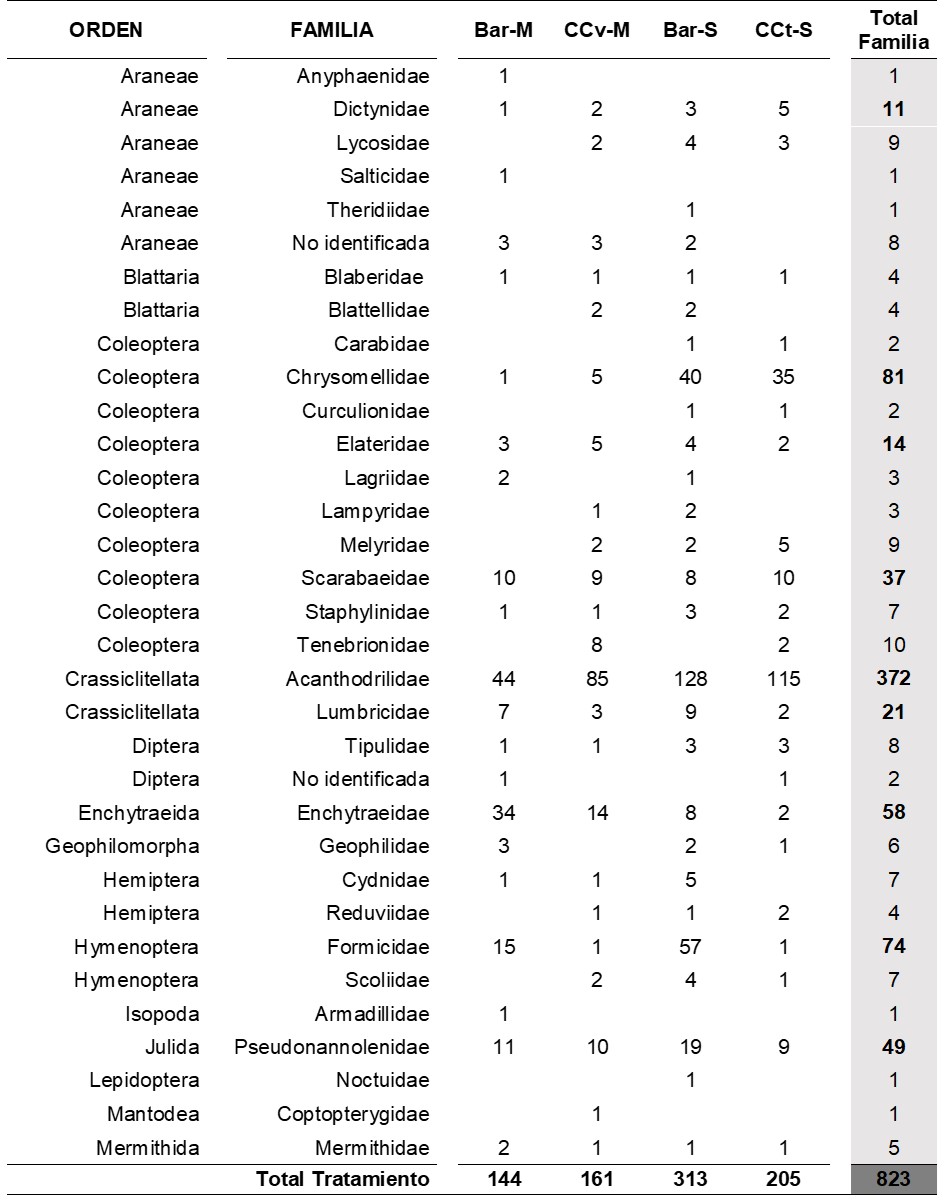
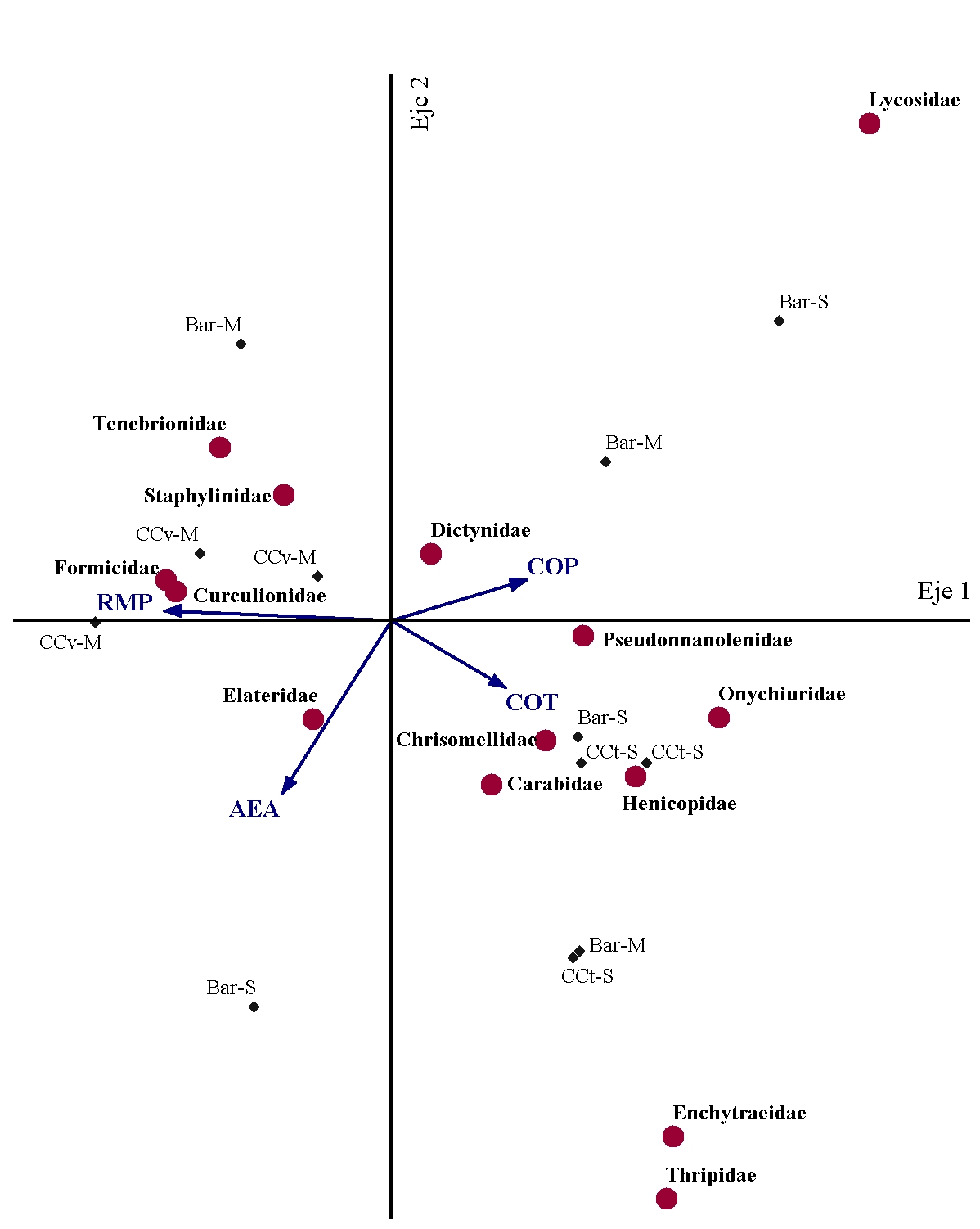
FIGURA 1/FIGURE 1
Figura 1. Análisis de Correspondencia Canónica entre las propiedades edáficas evaluadas y la abundancia de las familias de macrofauna más representativas en los diferentes tratamientos en el muestreo post CC y barbecho. Referencias: COT: carbono orgánico total; COP: carbono orgánico particulado; AEA; agregados estables al agua; RMP: resistencia mecánica a la penetración; Bar-M: barbecho-maíz; CCv-M: CC vicia-maíz; Bar-S: barbecho-soja; CCt-S: CC triticale-soja. Figure 1. Canonical Correspondence Analysis between the evaluated soil properties and abundance of the most representative families of macrofauna in the different treatments in post CC and fallow sampling. References: COT: total organic carbon; COP: particulate organic carbon; AEA: water stable aggregates; RMP: mechanical penetration resistance; Bar-M: fallow-corn; CCv-M: CC vetch-corn; Bar-S: fallow-soybean; CCt-S: CC triticale-soybean.
Las muestras con barbecho no presentaron un ordenamiento claro en el Mcc-b, pero se observó una vinculación de algunas de ellas con arañas de las familias Dictynidae y Lycosidae en asociación con bajos porcentajes de AEA que, en función de la información disponible, reflejaría cierto grado de degradación del suelo debido a este tipo de manejo. En este caso, dado que se trata de organismos con actividad epigea, los mismos podrían haberse favorecido frente a un menor volumen de materiales vegetales superficiales en tales situaciones que podrían obstaculizar su movimiento. En relación a ello, las arañas de la familia Lycosidae capturan a sus presas de manera activa, es decir, movilizándose en la superficie del suelo (Weeks y Holtzer, 2000) y han sido asociadas a sitios con alta intensidad de disturbio (Argañaraz et al., 2020). Por el contrario, las arañas de la familia Dictynidae construyen redes tipo “sábana” como estrategia para capturar a sus presas (Sharma et al., 2021), con lo cual podrían haberse visto favorecidas por la presencia de residuos del cultivo antecesor que servirían como anclaje para la elaboración de las redes.
En el Mf (Figura 2) se observó que las muestras correspondientes a los tratamientos que incluyeron CC se asociaron mayormente con valores superiores de COP e inferiores de RMP en relación a los de barbecho, en tanto que no se detectó un ordenamiento claro de las muestras en función del cultivo de cosecha. Asimismo, coincidentemente con lo observado en el Mcc-b, existió una mayor abundancia de hormigas en muestras con valores elevados de RMP y que se asociaron a las de Bar-S. Asimismo, se destacó que las lombrices de la familia Acanthodrilidae mostraron alta abundancia en muestras con elevado contenido de COP, entre las que se encontraron las correspondientes a las secuencias con CC, mientras que las de la familia Lumbricidae se asociaron con valores elevados de RMP. Tales respuestas revelarían diferencias en cuanto a las preferencias de hábitat de ambas familias de lombrices y, en el caso de Lumbricidae, la capacidad de desarrollarse en suelos con mayor dureza. Esto resulta relevante ya que se ha demostrado que las lombrices ejercen efectos positivos sobre numerosos aspectos del suelo, entre los que se destacan la contribución a la estructura edáfica, la retención de agua, la fertilidad y las comunidades microbianas, entre otros (Ahmed y Al-Mutairi, 2022; Al-Maliki et al., 2021). Al respecto, Bedano et al. (2019) indicaron que el incremento en la abundancia de lombrices en respuesta a una adecuada selección de prácticas de manejo promueve una mayor producción de agregados biogénicos, los cuales son más ricos en materia orgánica y más estables en comparación con el suelo circundante. Por otro lado, debe tenerse presente que la respuesta de estos organismos a las condiciones edáficas también podría variar en función de la especie considerada y su comportamiento en el suelo (Curry y Schmidt, 2007). En otro punto, las secuencias que incluyeron CC también se asociaron con las familias Scarabaeidae, Elateridae y Chrysomellidae, cuyos integrantes presentan hábitos herbívoros y fueron encontrados al estado larval. En estos casos, resulta interesante indicar que la condición físico-mecánica del suelo podría influir sobre la presencia de tales organismos ya que afectaría su capacidad de movilizarse para localizar las fuentes de recursos alimenticios (raíces) (Müller et al., 2022; Traugott et al., 2015). En relación a los resultados obtenidos, esto podría explicarse a partir de la asociación de las mencionadas familias con valores bajos de RMP.
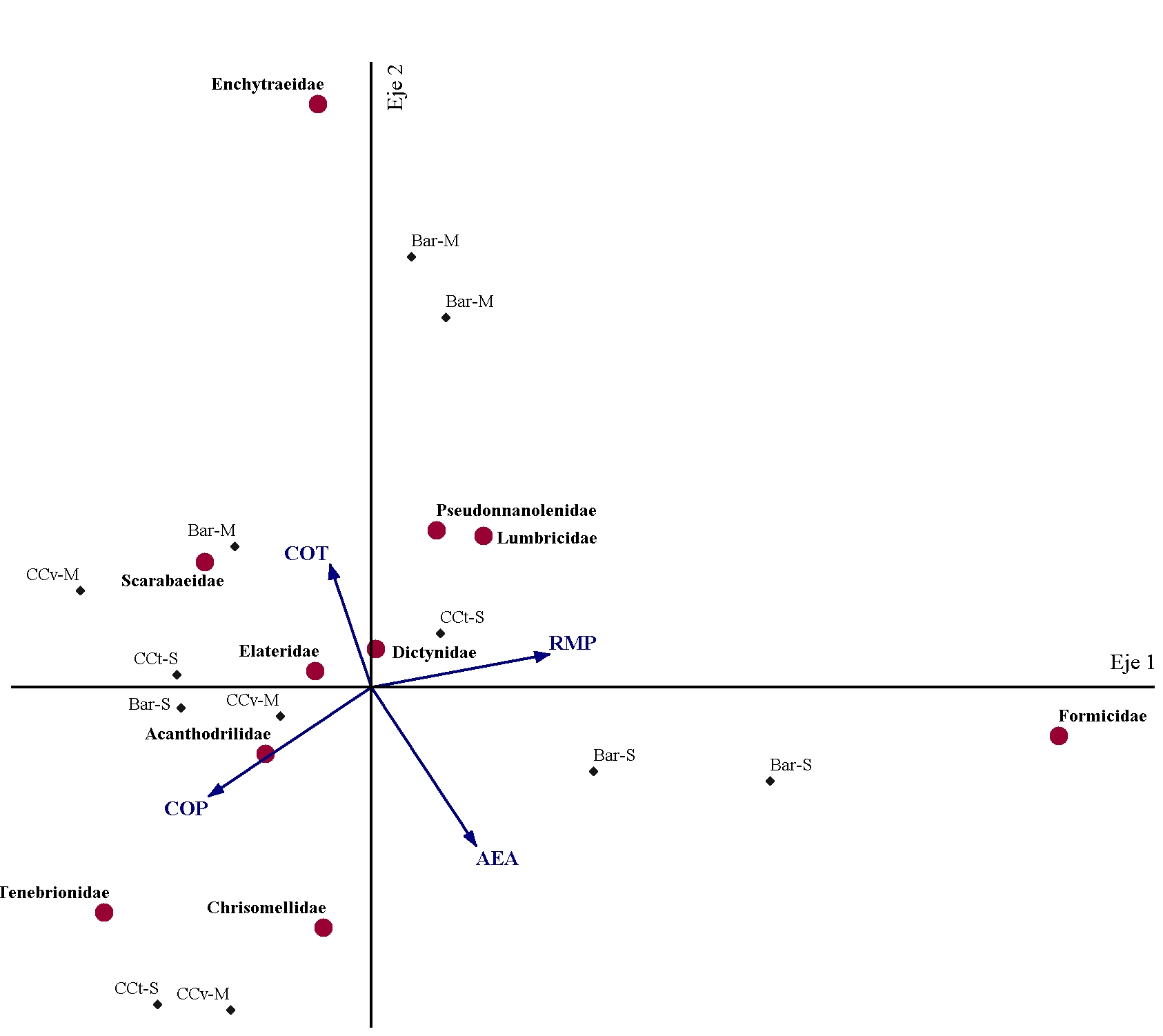
FIGURA 2/FIGURE 2
Figura 2. Análisis de Correspondencia Canónica entre las propiedades edáficas evaluadas y la abundancia de las familias de macrofauna más representativas en los diferentes tratamientos en el muestreo luego de la cosecha de los cultivos estivales. Referencias: COT: carbono orgánico total; COP: carbono orgánico particulado; AEA; agregados estables al agua; RMP: resistencia mecánica a la penetración; Bar-M: barbecho-maíz; CCv-M: CC vicia-maíz; Bar-S: barbecho-soja; CCt-S: CC triticale-soja. Figure 2. Canonical Correspondence Analysis between the evaluated soil properties and abundance of the most representative families of macrofauna in the different treatments in after cash crops harvest sampling. References: COT: total organic carbon; COP: particulate organic carbon; AEA: water stable aggregates; RMP: mechanical penetration resistance; Bar-M: fallow-corn; CCv-M: CC vetch-corn; Bar-S: fallow-soybean; CCt-S: CC triticale-soybean.
Conclusiones
La inclusión de CC en las secuencias de cultivos, principalmente de vicia, generó cambios a corto plazo en COP, RMP y, en menor medida, en el porcentaje de AEA, y que estuvieron asociados mayormente al aporte de residuos y el desarrollo radical durante sus ciclos de crecimiento. Asimismo, la abundancia de macrofauna también registró variaciones en el período de tiempo analizado, evidenciándose un incremento significativo luego del ciclo del CCv. Por otro lado, en los dos momentos evaluados existió una diferenciación de las muestras que incluyeron CC en relación a las de barbecho que permitió definir, aunque sea en relación a un conjunto determinado de propiedades, distinta condición del hábitat edáfico y, en base a ello, respuestas diferenciales de las principales familias de macrofauna.
En síntesis, los resultados obtenidos en este trabajo resaltan la importancia de incorporar CC en las secuencias de cultivo, teniendo en cuenta los cambios que podrían inducir en períodos de tiempo relativamente cortos. Además, se destaca la importancia de considerar a la macrofauna en la evaluación de las prácticas de manejo y la necesidad de profundizar en el estudio del hábitat edáfico y la respuesta de las comunidades de macroinvertebrados. Finalmente, sería acorde realizar investigaciones futuras a mediano/largo plazo y en condiciones reales de manejo, que vinculen tales tópicos con el rol de los CC en los cambios ambientales que éstos generan cuando se los incluye en las secuencias de cultivos.
Referencias
Ahmed, N., & Al-Mutairi, K. A. (2022). Earthworms effect on microbial population and soil fertility as well as their interaction with agricultural practices. Sustainability, 14, 7803. https://doi.org/10.3390/su14137803
Al-Maliki, S., Al-Taey, D. K. A., & Al-Mammori, H. Z. (2021). Earthworms and eco-consequences: considerations to soil biological and plant function: A review. Acta Ecologica Sinica, 41, 512-523. https://doi.org/10.1016/j.chnaes.2021.02.003
Anderson, J. M., & Ingram, J. S. I. (1993). Tropical soil biology and fertility. A handbook of methods (2da. Ed.). CAB International.
Argañaraz, C. I., Rubio, G. D., Rubio, M., & Castellarini, F. (2020). Ground-dwelling spiders in agroecosystems of the Dry Chaco: a rapid assessment of community shifts in response to land use changes. Biodiversity, 21(3), 125-135. https://doi.org/10.1080/14888386.2020.1831605
Bedano, J. C., Domínguez, A., Arolfo, R., & Wall, L. C. (2016). Effect of good agricultural practices under no-till on litter and soil invertebrates in areas with and without soil types. Soil and Tillage Research, 158, 100-109. http://doi.org/10.1016/j.still.2015.12.005
Bedano, J. C., Vaquero, F., Domínguez, A., Rodríguez, M. P., Wall, L., & Lavelle, P. (2019). Earthworms contribute to ecosystem process in no-till systems with high crop rotation intensity in Argentina. Acta Oecologica, 98, 14–24. https://doi.org/10.1016/j.actao.2019.05.003
Berg, G., & Smalla, K. (2009). Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiology Ecology, 68, 1–13. https://doi.org/10.1111/j.1574-6941.2009.00654.x
Blanco-Canqui, H., & Jasa, P. J. (2019). Do grass and legume cover crops improve soil properties in the long term? Soil & Water Management & Conservation, 83, 1181-1187. https://doi.org/10.2136/sssaj2019.02.0055
Blanco-Canqui, H., & Ruis, S. J. (2020). Cover crops impacts on soil physical properties: a review. Soil Science Society of America Journal, 84, 1527-1576. https://doi.org/10.1002/saj2.20129
Bodner, G., Leitner, D., & Kaul, H. P. (2014). Coarse and fine root plants affect pore size distributions differently. Plant and Soil, 380, 133-151. https://doi.org/10.1007/s11104-014-2079-8
Borror, D. J., Triplehorn, C. A., & Johnson, N. F. (1992). An introduction to the study of insects. Sunders College Publishing.
Cabrera, G. (2012). La macrofauna edáfica como indicador biológico del estado de conservación/perturbación del suelo. Resultados obtenidos en Cuba. Pastos y Forrajes, 35(4), 349-364.
Cambardella, C., & Elliot, E. (1992). Particulate soil organic matter changes across a grassland cultivation sequence. Soil Science Society of America Journal, 56, 777-783. https://doi.org/10.2136/sssaj1992.03615995005600030017x
Castiglioni, M., & Behrends Kraemer, F. (2019). Short-term effect of cover crops on aggregate stability assessed by two techniques. Ciencia del Suelo, 37(2), 298-314. http://www.suelos.org.ar/publicaciones/Volumen37n2/9-Pag%20298-314%20%23452.pdf
Chahal, I., Vyn, R. J., Mayers, D., & Van Eerd, L. L. (2020). Cumulative impact of cover crops on soil carbon sequestration and profitability in a temperate humid climate. Scientific Reports, 10, 13381. https://doi.org/10.1038/s41598-020-70224-6
Chanatásig-Vaca, C. I., Huerta, E., Rojas, P., Ponce-Mendoza, A., Mendoza, J., Morón, A., van der Wal, H., & Dzib-Castillo, B. B. (2011). Efecto del uso de suelo en las hormigas (Formicidae: Hymenoptera) de Tikinmul, Campeche, México. Acta Zoológica Mexicana, 27, 41-61. https://www.scielo.org.mx/pdf/azm/v27n2/v27n2a16.pdf
Chatterjee, A., Lal, R., Wielopolski, L., Martin, M. Z., & Ebinger, M. H. (2009). Evaluation of different soil carbon determination methods. Critical Reviews in Plant Science, 28, 164-178. https://doi.org/10.1080/07352680902776556
Crotty, F. V., & Stoate, C. (2019). The legacy of cover crops on the soil habitat and ecosystem services in a heavy clay minimum tillage rotation. Food and Energy Security, 8, e00169. https://doi.org/10.1002/fes3.169
Curry, J. P., & Schmidt, O. (2007). The feeding ecology of earthworms – A review. Pedobiologia, 50, 463-477. https://doi.org/10.1016/j.pedobi.2006.09.001
Di Rienzo, J. A., Macchiavelli, R. E., & Casanoves, F. (2011). Modelos lineales mixtos. Aplicaciones en InfoStat (1a. Ed.). Grupo InfoStat, UNC. https://www.researchgate.net/publication/283491350_Modelos_lineales_mixtos_aplicaciones_en_InfoStat
Di Rienzo, J. A., Macchiavelli, R. E., & Casanoves, F. (2017). Modelos lineales generalizados mixtos. Aplicaciones en InfoStat (1a. Ed.). Grupo InfoStat, UNC. https://www.researchgate.net/profile/FernandoCasanoves/publication/318768958_Modelos_lineales_generalizados_mixtos_aplicaciones_en_InfoStat/links/597cf833458515687b44eda1/Modelos-lineales-generalizados-mixtos-aplicaciones-en-InfoStat.pdf
Di Rienzo, J. A., Casanoves, F., Balzarini, M. G., González, L., Tablada, M., & Robledo, C. W. (2020). InfoStat (Versión 2020). Córdoba: Centro de Transferencia InfoStat. https://www.infostat.com.ar/
Duval, M. E., Galantini, J. A., Capurro, J. E., & Martínez, J. M. (2016). Winter cover crops in soybean monoculture: effects on soil organic carbon and its fractions. Soil & Tillage Research, 161, 95-105. http://dx.doi.org/10.1016/j.still.2016.04.006
Erktan, A., Or, D., & Scheu, S. (2020). The physical structure of soil: determinant and consequence of trophic interactions. Soil Biology and Biochemistry, 148, 107876. https://doi.org/10.1016/j.soilbio.2020.107876
Food and Agriculture Organization of the United Nations (FAO) (2020). State of knowledge of soil biodiversity. Status, challenges and potentialities. FAO. https://www.fao.org/documents/card/en/c/cb1928en
Fiorini, A., Remelli, S., Boselli, R., Mantovi, P., Ardenti, F., Trevisan, M., Menta, C., & Tabaglio, V. (2022). Driving crop yield, soil organic C pools and soil biodiversity with selected winter cover crops under no-till. Soil and Tillage Research, 217, 105283. https://doi.org/10.1016/j.still.2021.105283
Geisen, S., Briones, M. J. I., Gan, H., Behan-Pelletier, V. M., Friman, V. P., de Groot, G. A., Hannula, S. E., Lindo, Z., Philippot, L., Tiunov, A. V., & Wall, D. H. (2019). A methodological framework to embrace soil biodiversity. Soil Biology and Biochemistry, 136, 107536. https://doi.org/10.1016/j.soilbio.2019.107536
Halliday, W. D., Bourque, C., & Blouin-Demers, G. (2019). Food quality influences density-dependent fitness, but not always density-dependent habitat selection, in red flour beetles (Coleoptera: Tenebrionidae). The Canadian Entomologist, 151, 728-737. https://doi.org/10.4039/tce.2019.47
Hénin, S., Monnier, G., & Combeau, A. (1958). Méthode pour l´étude de la stabilité structurale des sols. Annales Agronomiques, 9, 73-92.
Hudek, C., Putinica, C., Otten, W., & De Baets, S. (2022). Functional root trait-based classification of cover crops to improve soil physical properties. European Journal of Soil Science, 73, e13147. https://doi.org/10.1111/ejss.13147
Instituto Nacional de Tecnología Agropecuaria (INTA). (13 de enero de 2023). Visor GeoINTA. http://visor.geointa.inta.gob.ar/
Jian, J., Du, X., Reiter, M. S., & Stewart, R. D. (2020). A meta-analysis of global cropland soil carbon changes due to cover cropping. Soil Biology & Biochemistry, 143, 107735. https://doi.org/10.1016/j.soilbio.2020.107735
Jiang, Y., Ma, N., Chen, Z., & Xie, H. (2018). Soil macrofauna assemblage composition and functional groups in no-tillage with corn stover mulch agroecosystems in a mollisol area of northeastern China. Applied Soil Ecology, 128, 61-70. https://doi.org/10.1016/j.apsoil.2018.04.006
Kelly, C., Fonte, S. J., Shrestha, A., Daane, K. M., & Mitchell, J. P. (2021). Winter cover crops and no-till promote soil macrofauna communities in irrigated, Mediterranean cropland in California, USA. Applied Soil Ecology, 166, 104068. https://doi.org/10.1016/j.apsoil.2021.104068
Krüger, H., Frolla, F. & Zilio, J. (2018). Un indicador de compactación relacionado con el agua del suelo. En A. Quiroga, R. Fernández & C. Álvarez (Eds.), Análisis y evaluación de propiedades físico hídrica de los suelos (1ra. ed. pp. 45-50). Ediciones INTA. https://ri.conicet.gov.ar/bitstream/handle/11336/114961/CONICET_Digital_Nro.004feb64-8f54-4a1a-8200-c2913b702596_B.pdf?sequence=5&isAllowed=y
Lavelle, P., Mathieu, J., Spain, A., Brown, G., Fragoso, C., Lapied, E., De Aquino, A., Barois, I., Barros, M. E., Bedano, J. C., Blanchart, E., Caufield, M., Chagueza, Y., Dai, J., Decaëns, T., Domínguez, A., Domínguez, Y., Feijoo, A., et al. (2022). Soil macroinvertebrate communities: A world-wide assessment. Global Ecology and Biogeography, 31, 1261-1276. https://doi.org/10.1111/geb.13492
Laossi, K. R., Barot, S., Carvalho, D., Dejardins, T., Lavelle, P., Martins, M., Rendeiro, A. C., Rousseau, G., Sarrazin, M., Velasquez, E., & Grimaldi, M. (2008). Effects of plant diversity on plant biomass production and soil macrofauna in Amazonian pastures. Pedobiologia, 51, 397-407. https://doi.org/10.1016/j.pedobi.2007.11.001
Masters, G. J. (2004). Belowground herbivores and ecosystem processes. En W. W. Weisser & E. Siemann (Eds.), Insects and ecosystem function (1ra. ed. pp. 93-109). Springer.
Mc Cune, B., & Mefford, M. J. (2011). PC-ORD. Multivariate Analysis of Ecological Data (Versión 6.0). Gleneden Beach, Oregon: MjM Software.
Menta, C., Conti, F. D., Lozano Fondón, C., Staffilani, F., & Remelli, S. (2020). Soil arthropod responses in agroecosystem implications of different management and cropping systems. Agronomy, 10, 982. https://doi.org/10.3390/agronomy10070982
Miransari, M. (2014). Plant growth promoting Rhizobacteria. Journal of Plant Nutrition, 37, 2227–2235. https://doi.org/10.1080/01904167.2014.920384
Morrone, J. J., & Coscarón, S. (1998). Biodiversidad de artrópodos argentinos (Vol. 1). Ediciones Sur. https://seargentina.com.ar/libros-digitales-de-libre-acceso/
Müller, P., Neuhoff, D., Nabel, M., Schiffers, K., & Döring, T. F. (2022). Tillage effects on ground beetles in temperate climates: a review. Agronomy for Sustainable Development, 42, 65. https://doi.org/10.1007/s13593-022-00803-6
Murrell, E. G. (2017). Can agricultural practices that mitigate or improve crop resilience to climate change also manage crop pests? Current Opinion in Insect Science, 23, 81-88. https://doi.org/10.1016/j.cois.2017.07.008
Roig-Juñent, S., Claps, L. E., & Morrone, J. J. (2014a). Biodiversidad de artrópodos argentinos (1ra. ed., Vol. 3). Sociedad Entomológica Argentina. https://seargentina.com.ar/libros-digitales-de-libre-acceso/
Roig-Juñent, S., Claps, L. E., & Morrone, J. J. (2014b). Biodiversidad de artrópodos argentinos (1ra. ed., Vol. 4). Sociedad Entomológica Argentina. https://seargentina.com.ar/libros-digitales-de-libre-acceso/
Romaniuk, R., Navarro, R., Beltrán, M., Eiza, M., Castiglioni, M., & Mousegne, F. (2018). Efecto a corto plazo de la inclusión de vicia y trigo como cultivos de cobertura sobre le C, N y P en distintas fracciones de la materia orgánica, y la disponibilidad de macro y micronutrientes. RIA, 44(2), 48-60. http://www.scielo.org.ar/pdf/ria/v44n2/v44n2a08.pdf
Rosario-Lebron, A., Leslie, A. W., Chen, G., & Hooks, C. R. R. (2018). The effect of barley cover crop residue and herbicide management on the foliar arthropod community in no-till systems. Agronomy, 8, 87. https://doi.org/10.3390/agronomy8060087
Sharma, A., Singh, G., & Singh, R. (2021). Faunal diversity of spider families Dictynidae, Dysderidae, Eresidae and Filistatidae (Araneomorphae: Araneae: Arachnida) in India. International Journal of Zoology and Applied Biosciences, 6(1), 1-9. https://doi.org/10.5281/zenodo.4460086
Stasiov, S., Vician, V., Bencat, T., Pätoprstý, V., Lukácik, I., & Svitok, M. (2021). Influence of soil properties on milipede (Diplopoda) communities in forest stands of various tree species. Acta Oecologica, 113, 103793. https://doi.org/10.1016/j.actao.2021.103793
Stehr, F. W. (1991). Immature insects (1ra. ed., Vol. 2). Kendall-Hunt Publishing Company.
Sylvain, Z. A., & Wall, D. H. (2011). Linking soil biodiversity and vegetation: implications for a changing planet. American Journal of Botany, 98(3), 517-527. http://www.jstor.org/stable/41149201
Ter Braak, C. J. F. (1986). Canonical Correspondence Analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology, 67, 1167-1179.
Traugott, M., Benefer, C. M., Blackshaw, R. P., van Herk, W. G., & Vernon, R. S. (2015). Biology, ecology, and control of elaterid beetles in agricultural land. Annual Review of Entomology, 60, 313–334. https://doi.org/10.1146/annurev-ento-010814-021035
Wagg, C., Bender, S. F., Widmer, F., & van der Heijden, M. G. A. (2014). Soil biodiversity and soil community composition determine ecosystem multifunctionality. PNAS, 111, 5266-5270. https://doi.org/10.1073/pnas.1320054111
Walkley, A., & Black, I. A. (1934). An examination of Degtjareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Science, 37, 29-37.
Weeks, R. D., & Holtzer, T. O. (2000). Habitat and season in structuring ground-dwelling spider (Araneae) communities in a short grass steppe ecosystem. Environmental Entomology, 29(6), 1164-1172.
White, C. A., Holmes, H. F., Morres, N. L., & Stobart, R. M. (2016). A review of the benefits, optimal crop management practices and knowledge gaps associated with different cover crop species. Research Review, 90. AHDB. https://ahdb.org.uk/a-review-of-the-benefits-optimal-crop-management-practices-and-knowledge-gaps-associated-with-different-cover-crop-species
Zhang, Z., Kaye, J. P., Bradley, B. A., Amsili, J. P., & Suseela, V. (2022). Cover crop functional types differentially alter the content and composition of soil organic carbon in particulate and mineral associated fractions. Global Change Biology, 28, 5831-5848. https://doi.org/10.1111/gcb.16296
Notas de autor
scaglionejosefina@gmail.com

